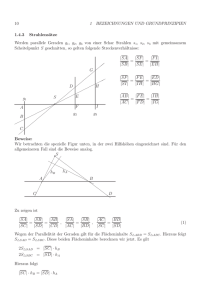
Dynamik Neuronaler Netze
Werbung

Dynamik Neuronaler Netze
Ausarbeitung von Ercan Elitok nach „Theoretical Neuroscience” von Peter Dayan, L. F. Abbott, p. 260273
Inhalt
Dynamik Neuronaler Netze........................................................................................................ 1
Netzwerkstabilität ................................................................................................................... 2
Assoziatives Gedächtnis/Speicher......................................................................................... 3
Exzitatorisch-Inhibitorische Netzwerke .................................................................................. 7
Homogene Exzitatorische ond Inhibitorische Populationen .............................................. 8
Der Bulbus olfaktorius ...................................................................................................... 11
Oszillatorische Verstärkung ............................................................................................. 14
Legende
τ s = Zeitkonstante bis Erreichen des stationären Zustands des Neurons S,
v = F ( I s ) = Netzwerkfeuerrate, mit I s als Vektor des gesamten synaptischen Stroms
des Neurons S,
u = Inputvektor,
v = Outputvektor,
h = gesamter vorwärts gekoppelter Input,
w = Inputgewichtsvektor,
W = Gewichtsmatrix des Inputs,
M = Gewichtsmatrix des Outputs bzw. der Rückwärtskopplung,
t = Zeit.
Zusammenfassung
Die Dynamik neuronaler Netze kann mit unterschiedlichen Techniken analysiert
werden. Mit der Theorie der Lyapunovfunktionen kann untersucht werden, ob ein
Netzwerk Fixpunktverhalten zeigt. Hierbei wird deutlich, dass die Gewichtsmatrizen
eines neuronalen Netzwerks Stabilitätskriterien erfüllen müssen, die sich von eine r
Lyapunovfunktion ableiten. Am Beispiel eines assoziativen Speichernetzwerks wird
deutlich, dass zur Herleitung eines Modells die Eigenschaften der zugehörigen
symmetrischen Gewichtsmatrix wesentlich durch die zu bewerkstelligenden
Aufgaben des Netzwerks bestimmt sind. Das Netzwerkmodell muss zum Auslesen
bestimmter Muster passende Fixpunkte besitzen. Modelle exzitatorischinhibitorischer Neuronenpopulationen, die eine asymmetrische Gewichtsmatrix
besitzen zeigen neben einem Fixpunktverhalten auch oszillierende Aktivität. Die
Beurteilung, ob es sich um stabile bzw. instabile Fixpunkte oder um oszillatorisches
Netzwerkverhalten handelt kann mittels Phasenebenen- und Stabilitätsanalyse
untersucht werden. Hierbei sind die (komplexen) Eigenwerte der Gewichtsmatrix die
entscheidenden Kriterien für die Analyse. Ein Beispiel für ein exzitatorischinhibitorisches Modell stellt die Simulation des Bulbus olfaktorius dar. Hier können
Modell- und biologische Daten miteinander verglichen werden. Ein Beispiel für
oszillierende Aktivität eines exzitatorich-inhibitorischen Netzwerkmodells findet sich
bei auf bestimmten Input spezialisierten Neuronenpopulationen, deren Aktivität durch
abgestimmten Input selektiv verstärkt werden kann.
1
Netzwerkstabilität
Wenn ein Netzwerk bei einem konsta nten Input zu einem stationären Zustand
übergeht mit dv/dt=0, dann spricht man davon, dass das Netzwerk ein FixpunktVerhalten (fixed-point behavior) zeigt. Dies ist zwar nicht die einzige Form einer
Langzeitaktivität, die ein Netzwerkmodel annehmen kann, aber beim Vorliegen
bestimmter Bedingungen erreicht ein Netzwerk als Antwort auf einen konstanten
Input unausweichlich einen Fixpunkt. Die Theorie der Lyapunovfunktionen kann dazu
benutzt werden um dieses Netzwerkverhalten nachzuweisen was im Folgenden
einführend informal beschrieben wird.
Zur Diskussion der Lyapunovfuktionen betrachten wir zunächst die Dynamik der
Feuerrate
τs
dI s
= − I s + w ⋅ u , mit v = F ( I s ) ,
dt
(1)
In einem Netzwerkmodell stellt v = F (I ) den Vektor der Feuerrate dar, mit
F ' ( I ) > 0, ∀ I . I befolgt die Dynamik der Formel
dI
τs
= − I + h + M ⋅ F (I ) ,
dt
(2)
die die verallgemeinerte Form der Gleichung 1 für ein Netzwerkmodell darstellt.
Die Gleichung 2 kann auch dafür verwendet werden um das Feuerratenmodell von
rückwärtsgekoppelten Netzwerken darzustellen.
Cohen und Grossberg (1983) haben für diese Form des Feuerratenmodells
(Gleichung 2 mit einer symmetrischen rückwärtsgekoppelten Gewichtsmatrix M )
gezeigt, dass durch
N
Ia
1 v
L( I ) = ∑ ∫ dz a z a F ' ( z a ) − ha F ( I a ) − ∑ F ( I a ) M aa' F ( I a ' )
2 a '=1
a =1 0
Nv
eine Lyapunovfunktion gegeben ist. Um das nachzuvollziehen setze man die
dL
Ableitung der Gleichung 3
in die Gleichung 2 ein. Dies ergibt:
dt
2
dL( I )
1 Nv
dI a
= − ∑ F ' ( I a )
dt
τ s a=1
dt
(3)
(4)
Es gilt: dL / dt < 0 solange dI / dt ≠ 0 und F ' > 0. Ferner ist L ist nach unten beschränkt
und kann somit nicht beliebig kleiner werden. Daher muss I = h + M ⋅ v zu einem
Fixpunkt konvergieren. Hieraus folgt, dass v = F (I ) ebenfalls zu einem Fixpunkt
konvergieren muss.
Wir haben hierfür vorausgesetzt, dass F ' ( I ) > 0 , ∀I . Jedoch können wir zeigen,
dass die oben gezeigte Lyapunovf unktion ebenfalls gültig ist für den Fall der
Rektifizierung der Aktivierungsfunktion F ( I ) = [I ]+ , mit [I ]+ = max{ 0, I } . Weiterhin ist
es notwendig zu zeigen, dass die Lyapunovfunktion eine untere Schranke hat. Eine
Möglichkeit wäre eine sättigende Aktivierungsfunktion F zu benutzen, die bei
2
I → ∞ eine Schranke F * = lim F ( I ) hat und eine zweite Möglichkeit wäre die
I→∞
Gewichtsmatrix so zu wählen, dass ihre Eigenwerte genügend klein sind.
Assoziativer Speicher
Die Netzwerkmodelle für Gedächtnis, die mittels anhaltender Aktivität Informationen
speichern werden Arbeits- bzw. Kurzzeit-Gedächtnis gena nnt. In biologischen
Systemen scheint es so zu sein, dass die anhaltende Aktivität dazu dient
Informationen über einen Zeitraum von Sekunden bis Minuten zu speichern. Die
Informationsspeicherung im Langzeitgedächtnis, d.h. Speicherung über einen
Zeitraum von Stunden bis Jahren, scheint mehr über die Stärke der synaptischen
Verbindung als über anhaltende Aktivität zu erfolgen. Eine allgemeine Idee ist die
synaptische Gewicht in einem rückwärtsgekoppelten Netzwerk beim
Speichervorgang eines Gedächtnisinhaltes so zu setzen, dass das Netzwerk zu
einem späteren Zeitpunkt von sich heraus das Aktivitätsmuster des Speicherinhaltes
wiederherstellen kann. In solchen Netzwerken wird die persistente Aktivität sowohl
zur Signalisierung des Speicheraufrufs als auch zum Anzeigen der Identität des
aufgerufenen Speicherinhaltes benutzt und nicht für die Langzeitspeicherung der
möglichen Speichermuster. Das Aktivitätsmuster der Netzwerkneuronen bestimmt zu
Beginn des Speicheraufrufs welcher Informationsinhalt aufgerufen wird und zwar
vermittelt durch die Beziehung bzw. Assoziation des Aktivitätsmusters mit dem
Muster des Speicherinhalts.
Diese assoziativen Netzwerke wurden benutzt, um Gehirnregionen von Säugern mit
unterschiedlichen Gedächtnisformen einschließlich der Area CA3 des Hippokampus
und des präfrontalen Kortex zu modellieren.
In einem assoziativen, genauer autoassoziativen Speicher, wird ein Teil oder eine
annähernd gleiche Repräsentation eines gespeicherten Elements dazu benutzt um
das ganze Element abzurufen. Anders als bei einem Computer basiert in einem
assoziativen Gedächtnis der Speicherabruf mehr auf dem Inhalt als auf der Adresse
der Speicherstelle. Beispielsweise würde in einem Netzwerkmodell eines
assoziativen Speichers eine komplette Telefonnummer dadurch aufgerufen, dass
anfänglich nur einige Ziffern der Nummer als Anhaltspunkt angegeben werden. In
solchen Netzwerken sind die rückwärtsgekoppelten synaptischen Gewichte so
eingestellt, dass das Netzwerk ein Menge von diskreten Fixpunkten hat, die (fast)
identisch mit dem Speicherinhaltrepräsentierenden Aktivitätsmuster sind. In vielen
Fällen wird die Dynamik des Netzwerks durch die Lyapunovfunktion (Gleichung 2)
bestimmt, vorausgesetzt Fixpunkte existieren. Um nicht unnötig viele Informationen
zu speichern, entsprechen die Fixpunkte genau oder zumindest annähernd genau
dem Muster der Informationen. Während eines Speicheraufrufs wird in einem
assoziativen Gedächtnisnetzwerk die Mustererkennung derart berechnet, dass die
Fixpunkte gefunden werden die dem Anfangszustand des Netzwerkes am
ähnlichsten sind. Jedes Speichermuster hat einen Einzugsbereich (basin of
attraction), welcher durch die Menge der anfänglichen Aktivitätsmuster, die zu dem
jeweiligen Fixpunkt konvergieren, definiert ist. Dieser Einzugsbereich bestimmt die
Ausleseeigenschaften des Netzwerks.
3
Assoziative Gedächtnisnetzwerke könne durch Neuronen hergestellt werden, deren
Aktivität entweder mit stetigen oder mit binären (on/off) Werten charakterisiert wird.
Wir betrachten ein Netzwerk mit Neuronen stetiger Aktivitätswerte welches durch
τr
dv s
= −v + F ( h + M ⋅ v) mit h = 0 , ( h = W ⋅ u)
dt
(5)
beschrieben wird. Um dieses Modell für die Informationsspeicherung nutzen zu
können, definieren wir eine Mustermenge mit der Bezeichnung v m , ( m = 1,2,..., N mem ),
der sowohl für die Speicherung als auch für den Speicheraufruf genutzt wird. Hierbei
stellt v m den Vektor mit der Nummer m dar. Ein assoziativer Speicheraufruf erfolgt
indem das Netzwerk in einem Anfangszustand gestartet wird, der einem der
Informationsmuster ähnlich ist. D.h. v (0) ≈ v m für ein m, wobei das „ ≈" bedeutet, dass
der Startwert v (0) in der Nähe von v m liegt. Das Netzwerk verhält sich entsprechend
der Gleichung 5. Wenn der Datenaufruf erfolgreich ist, dann konve rgiert die Dynamik
des Netzwerks zu einem Fixpunkt, der dem Informationsmuster gleich oder
zumindest bedeutend ähnlicher als der Anfangsstatus v (0) ist, d.h. v (t ) → v m für
große t . Ein Datenaufruf misslingt entweder wenn der erreichte Fixpunkt v m nicht
gleicht oder ein Fixpunkt erst gar nicht erreicht wird.
Für einen genauen Datenaufruf ist es notwendig, dass v m ein Fixpunkt der
Netzwerkdynamik ist bzw.
v m = F (M ⋅ v m )
(6)
erfüllt.
Die Kapazität eines Netzwerkes ist beispielsweise durch die Anzahl verschiedener
Vektoren bestimmt, die bei geeigneter Wahl von M gleichzeitig Gleichung 6 erfüllen.
Bei großen N v (= Anzahl Neuronen im Netzwerk) ist die Netzwerkkapazität
typischerweise proportional zu N v . Neben der Kapazität des Netzwerks ist auch die
Anzahl der Fixpunkte für die Leistungsfähigkeit eines assoziativen
Speichernetzwerks von Bedeutung. Die Leistungsfähigkeit des Netzwerks wird durch
Fixpunkte, die nicht durch Speichermuster entstanden sind, herabgesetzt. Für
effiziente Musterabgleichung ist für jeden der Fixpunkte ein geeignet große r
Einzugsbereich notwendig.
Im Folgenden gehen wir von Neuronen im Netzwerk aus, die sich binär verhalten
(aktiv/inaktiv) obwohl sie stetige Werte des Aktivitätszustandes liefern könnten.
Inaktive Neuronen entsprechen Nullen und aktive Neuronen einer Konstanten c in
den Komponenten von v m . Zur Vereinfachung nehmen wir an, dass das Muster
genau α N v aktive und (1 − α ) N v inaktive Neuronen besitzt. Die Auswahl der aktiven
Neuronen ist zufällig und unabhängig von anderen Mustern. Hierbei stellt α die
Spärlichkeit (sparseness) dar. Je kleiner α ist, desto spärlicher sind die Muster.
Hierdurch können zwar mehr Muster gespeichert werden, jedoch nur auf Kosten
eines geringeren Informationsgehaltes pro Muster.
Um ein assoziatives Speichernetzwerk herzustellen konstruieren wir eine Matrix so,
dass die Informationsmuster die Gleichung 6 erfüllen. Zunächst gehen wir davon aus
4
eine Matrix K zu kennen, für die alle Muster degenerierte Eigenvektoren mit dem
Eigenwert λ darstellen,
K ⋅ v m = λv m , ∀m .
(7)
Danach betrachte n wir die Matrix
M =K−
nn
1
oder M aa ' = K aa' −
αN v
αN v
(8)
Hierbei ist n = (1,1,...,1) der 1-Vektor mit N v Einsen. Der Term nn stellt die
gleichförmige Inhibition zwischen Netzwerkneuronen dar. M erfüllt für jedes
Informationsmuster v m die
M ⋅ v m = λv m − cn .
(9)
Die rechte Seite des Terms folgt aus der Tatsache, dass n ⋅ v m = cαN v ist. Bearbeitet
man die Gleichung 6 komponentenweise für diese Matrix, so bilden sich zwei
Bedingungen: die erste für die Komponenten i mit v im = 0 die Null und die andere für
die Komponenten i von v im = c sind,
F ( −c ) = 0 und c = F (c (λ − 1)) .
(10)
Es ist vergleichsweise einfach Bedingungen zu finden, für die diese Gleichungen
eine Lösung haben. Für positive c ist die erste Bedingung, mit einer rektifizierende
Aktivierungsfunktion F ( I ) = 0 , mit I ≤ 0 , automatisch erfüllt. Für alle F ' ( I ) = 0 , mit
positiven I wird die zweite Gleichung (von Gleichung 10) erfüllt und die Gleichung 5
würde für c > 0 einen stabilen Fixpunkt als Lösung haben, wenn beispielsweise
λ > 1, ( λ − 1) F ' (0) > 1 und F ( c( λ − 1)) langsamer wachsen als c (für große c ).
Das Problem ein assoziatives Speichernetzwerk zu konstruieren besteht darin, die
Matrix K aus Gleichung 7 oder zumindest eine mit ähnlichen Eigenschaften zu finden.
Sehen wir voraus, dass die Neuronen mit Wahrscheinlichkeit α aktiv sind, d.h. deren
Ausgabewert auf c gesetzt ist, so ist die Wahrscheinlichkeit, dass dasselbe Neuron
in zwei verschiedenen Informationsmuster n und m aktiv ist gleich α 2 . Daraus folgt,
dass v n ⋅ v m ≈ α 2 c 2 N v , für n ≠ m. Würde einem der beiden Informationsmuster
entsprechender Vektor v m mit dem Vektor v n − αcn mittels Skalarprodukt
multipliziert, so wäre für m = n ( v n − α cn) ⋅ v m = c 2αN v (1 − α ) und für m ≠ n wäre
( v n − αcn ) ⋅ v m ≈ c 2α N v (α 2 − α 2 ) = 0 . Hieraus folgt, dass die Matrix
K=
Nmem
λ
v n (v n − αcn)
∑
2
c αN v (1 − α ) n=1
(11)
ähnliche Eigenschaften hat wie die Matrix in Gleichung 7, d.h. K ⋅ v m ≈ λv m , ∀m .
5
Erinnern wir uns noch einmal daran, dass die Lyapunovfunktion in Gleichung 3 nur
dann zusichert, dass das Netzwerk Fixpunkte hat, wenn es eine untere Schranke hat
und die Matrix M symmetrisch ist. Die Beschränkung der Lyapunovfunktion kann
durch Sättigung der Aktivierungsfunktion erreicht werden. Die rückwärtsgekoppelte
Gewichtsmatrix K , die man durch das Einsetzen des Ausdrucks aus Gleichung 11 in
Gleichung 8 erhält, scheint nicht symmetrisch zu sein. Eine Form des Ausdrucks
kann hergestellt werden, in dem man
M i, j =
Nmem
λ
nn
(vin − αcn)( v nj − αcn ) −
∑
2
c αN v (1 − α ) n=1
αN v
(12)
verwendet. Durch das Hinzufügen dieser zusätzlichen Terme in den
Summenausdruck (über die Muster) sind die Bedingungen aus Gleichung 10, die für
die Nutzung von Gleichung 12 erfüllt sein müssen, verändert zu
F ( −c (1 + αλ )) = 0 und c = F (c (λ − 1 − αλ)) .
(13)
Eine Möglichkeit die rückwärtsgekoppelte Gewichtsmatrix in Gleichung 12 zu
betrachten ist die vom Standpunkt einer Lernregel, die dazu benutzt wurde um die
Matrix herzustellen. Bei dieser Lernregel wird zur Verbindung zweier Neuronen ein
exzitatorischer Beitrag immer dann hinzugefügt, we nn beide Neuronen bei einem
bestimmten Informationsmuster entweder aktiv oder inaktiv sind. Ein inhibitorischer
Ausdruck wird hinzugefügt, wenn ein Neuron aktiv und die andere inaktiv ist. Die mit
Gleichung 12 verbundene Lernregel wird wegen ihrer Beziehung zu der
Kovarianzmatrix der Informationsmuster als Kovarianzregel bezeichnet.
Abbildung 1: Aufruf eines Informationsmusters in einem assoziativen Speichernetzwerkmodell mit
50 Neuronen. (A) zeigt die Feurungsraten zweier repräsentativer Neuronen (oben Neuronaktivität
einer nicht-null- und unten Neuronaktivität einer null-Komponente des Informationsmusters). (B) und
(C) zeigen die Feurerungsraten aller 50 Neuronen, wobei die Dicke der Striche proportional zur
Feuerrate ist. In beiden Fällen konvergiert die Aktivität nach anfänglich zufälliger Aktivität der
Netzwerkneuronen zu dem jeweiligen Muster in (B) und C. Die Endmuster ähneln jeweils dem
gespeicherten Informationsmuster. Das Modell benutzt die Matrix aus Gleichung 12 mit α = 0.25,
λ = 1.25
und
F ( I ) = 150 Hz [tanh(( I + 20 Hz / 150Hz )) ]+ .
6
Die Abbildung 1 zeigt ein assoziatives Speichernetzwerk mit 50 = N v Neuronen unter
der Verwendung der Matrix aus Gleichung 12, das vier Muster speichert. Zwei dieser
Muster sind zufällig und um sie in der Abbildung leichter zu finden sind die beiden
anderen Muster sind nicht zufällig generiert. Ihr Aufruf ist in der Abbildung 1B und 1C
gezeigt. Von einem anfänglichen Muster der Aktivität gleicht es nur undeutlich einem
der gespeicherten Muster. Das Netzwerk erreicht einen Fixpunkt, der der
bestmöglichen Angleichung zu dem gespeicherten Informationsmuster sehr nahe
kommt. Wendet man die gleiche Prozedur auf die beiden anderen vom Netzwerk
gespeicherten Informationsmuster, so sind sie schwerer in der Abbildung zu
erkennen.
Die Matrix M von Gleichung 12, die wir als Basis für die Herleitung eines
assoziativen Speichernetzwerks benutzt haben, erfüllt die Bedingungen für genaue
Speicherung und das Auslesen von Informationsmustern nur annäherungsweise.
Diese Matrix führt einige Fehler in den Datenaufruf ein. Mit steigender Anzahl der zu
speichernden Muster verschlechtert sich die Approximation sowie die
Leistungsfähigkeit des assoziativen Speichers und senkt die Anzahl der
Informationen, die gespeichert werden können. Die einfache Beschreibung der
Kovarianz der Gewichte in Gleichung 12 ist weit vom Optimum entfernt. Andere
Vorschriften zur Erstellung von M können bedeutend höhere Speicherkapazitäten
ermöglichen.
Die Essenz der Untersuchung von assoziativen Gedächtnis Netzwerken ist, dass
große Netzwerke große Anzahl von Mustern speichern können insbesondere wenn
sie spärlich sind ( α klein ist) und beim Auslesen einige Fehler toleriert werden
können. Die Kapazität von bestimmten assoziativen Speichernetzwerken kann
analytisch berechnet werden. Die Anzahl der Informationsmuster, die gespeichert
werden können ist in der Größenordnung, wenngleich typischerweise kleiner als der
Neuronenanzahl in dem Netzwerk N v sowie abhängig von der Spärlichkeit
(sparseness) α . Die Menge der Information, die gespeichert werden kann ist
proportional zu N v2 , was annähernd der Anzahl der im Netzwerk befindlichen
Synapsen entspricht, wobei die gespeicherte Information je Synapse, d.h. die
Proportionalitätskonstante, typischerweise relativ klein ist.
Exzitatorisch-Inhibitorische Netzwerke
In diesem Abschnitt werden Modelle besprochen, in denen exzitatorische und
inhibitorische Neurone voneinander getrennt durch die
dv
τ E E = −v E + FE (hE + M EE ⋅ v E + M EI ⋅ v I ) und
(14)
dt
dv
τ I I = −v I + FI ( hI + M II ⋅ v I + M IE ⋅ v E )
(15)
dt
beschrieben werden. Diese Modelle sind dynamischer als die
Einzelpopulationsmodelle mit symmetrisch gekoppelten Matrizen. In Modellen mit
exzitatorisch-inhibitorischen Subpopulationen sind die synaptischen
Gewichtsmatrizen nicht symmetrisch, wodurch im Netzwerk Oszillationen entstehen
können. Wir analysieren nun ein Modell mit gleichförmig exzitatorisch-inhibitorischen
Populationen. Wir betrachten Methoden, die feststellen ob das Modell eine konstante
oder oszillierende Aktivität hervorbringt. Danach werden zwei Modelle besprochen, in
7
denen Oszillationen entstehen. Das erste ist ein Modell des Bulbus olfaktorius und
das zweite zeigt eine selektive Verstärkung in einem oszillierenden Modus.
Homogene Exzitatorische und Inhibitorische Populationen
Um die Dynamik in einem exzitatorisch-inhibitorischen Netzwerk zu veranschaulichen
analysieren wir ein einfaches Modell in welchem alle exzitatorischen Neurone durch
eine einzelne Feuerrate v E und die inhibitorischen mit einer zweiten Feuerrate v I
beschrieben werden. Obwohl wir uns dieses Modell als eine Interaktion zwischen
Neuronenpopulationen vorstellen, ist es so aufgebaut als ob es sich um nur zwei
Neurone handelt. Die Gleichung 14 und 15, zusammen mit den überschwelligen
linearen Antwortfunktionen, beschreiben diese zwei Feuerraten mit
τE
dv E
= −v E + [M EE v E + M EI v I − γ E ]+ und
dt
(16)
τI
dv I
= −v I + [M II v I + M IE v I − γ I ]+ .
dt
(17)
In diesem Modell sind die synaptische n Gewichte M EE , M IE , M EI und M II als Zahlen
und nicht als Matrizen zu verstehen. In diesem Beispiel seien M EE = 1.25, M IE = 1,
M EI = −1, M II = 0, γ E = −10Hz, γ I = 10Hz, τ E = 10 ms und τ I wird variiert. Der
negative Wert von γ E bedeutet hier nur, dass sie eine konstante Hintergrundaktivität
darstellt und keinen Schwellenwert.
Phase-Plane Methoden und Stabilitätsanalyse
Das durch die Gleichung 16 und 17 dargestellte Modell der Interaktion von
exzitatorisch und inhibitorischen Populationen bietet sich die Gelegenheit einige
Techniken, die zur Untersuchung der Dynamik der nichtlinearen Systeme nötig sind
zu veranschaulichen. Dieses Modell weist in Abhängigkeit von den Werten der
Parameter sowohl Fixpunkt- (konstantes v E und vI ) als auch oszillierende Aktivität
auf. Stabilitätsanalyse dient dazu um herauszufinden welche Parameterwerte zu
einem Übergang zwischen den beiden Formen der Aktivität führt.
Die Feuerraten v E (t ) und v I (t ) aus den Gleichungen 16 und 17 können als eine
Funktion der Zeit dargestellt werden, wie sie in Abbildung 3A und 4A zu sehen sind .
Eine andere nützliche Darstellung ist die paarweise Darstellung von v E (t ) und v I (t )
für ein Definitionsbereich von t -Werte n ( Abbildung 3B und 4B). Mit Änderung der
Feuerrate zeichnet sich eine Kurve bzw. Trajektorie in der v E - v I -Ebene ab, die als
die Phasenebene („phase plane“) des Modells bezeichnet wird. Die grafische
Darstellung von Phasenebenen gibt ein geometrisches Abbild der Dynamik eines
Modells wieder.
Von besonderem Interesse in der Phasenebenenanalyse sind die Werte für v E und
vI , für die die rechte Seite der Gleichungen 16 und 17 Null wird. Eine Reihe von
dv E
dv I
Werten für v E und vI (für die gilt
= 0 und
= 0 ), bilden zwei Linien in der
dt
dt
Phasenebene die als Nullkline bezeichnet werden. Die Nullklinen der Gleichungen 16
und 17 sind die geraden Linien in der Abbildung 2A. Die Nullklinen sind wichtig, weil
8
sie die Phasenebene in Gebiete einteilen mit gegensätzlichen Flussmustern. Das ist
dv E
dv I
so, weil wie in den Gleichungen 16 und 17 nachvollziehbar
und
auf der
dt
dt
einen Seite ihrer Nullkline positiv und auf der anderen negativ sind. Entlang der
dv E
Nullkline ist
gleich, oberhalb kleiner und unterhalb größer Null. Analog ist
dt
dv I
entlang der Nullkline
gleich, rechts kleiner und links größer Null. Diese Werte
dt
bestimmen die Flussrichtung in der Phasenebene und sind in der Abbildung 2A als
vertikale und horizontale Pfeile zu sehen. Typischerweise verringert sich die
Flussrate, wenn die Trajektorie der Phasenebene sich einer Nullkline nähert.
Abbildung 2: (A) Nullklinen, Flussrichtungen und Fixpunkt des Feuerratenmodells von
interagierenden exzitatorisch und inhibitorischen Neuronen. (B) der Real- (oben) und Imaginärteil
(unten) des Eigenwertes der die Stabilität des Fixpunkts bestimmt.
An einem Fixpunkt eines dynamischen Systems bleiben die Werte der
veränderlichen Variablen konstant. In diesem besprochenen Modell entsteht ein
dv E dv I
Fixpunkt wenn die Feuerraten v E und vI Werte annehmen für die
=
= 0 ist.
dt
dt
Dieser Fall tritt nur an der Schnittstelle der Nullklinen auf. In unserem Beispiel tritt
das bei v E = 26.67 und v I = 16.67 auf (aufgefüllter Kreis in der Mitte der Abbildung
2A). Ein Fixpunkt liefert zwar eine potentiell statische Konfiguration, jedoch ist es von
entscheidender Wichtigkeit ob der Fixpunkt stabil oder instabil ist. Wenn ein Fixpunkt
stabil ist werden die anfänglich in seiner Nähe befindlichen Werte von v E und vI über
die Zeit zu ihm hingezogen. Im Falle eines instabilen Fixpunktes wird das System bei
nur annähernd genauen Feuerraten von v E und vI vom Fixpunkt weggedrückt, es
sei denn die Feuerraten von v E und vI sind von Anfang an mit unendlicher
Genauigkeit auf die Werte des Fixpunktes gesetzt, denn dann bleibt das System
unbegrenzt im Fixpunkt.
Lineare Stabilitätsanalyse kann dazu benutzt werden um zu bestimmen, ob ein
Fixpunkt stabil ist oder nicht. Um diese Analyse durchzuführen benutzen wir
dv E
dv I
Abkömmlinge der Ausdrücke
und
, die dadurch entstehen indem die rechte
dt
dt
9
Seite der Gleichung 16 durch τ E und die der Gleichung 17 durch τ I dividiert wird.
Danach werden die dem Fixpunkt entsprechenden Werte für v E und vI in diese
Ausdrücke eingesetzt. Die auf diese Weise berechneten vier Terme werden dann in
einer Matrix angeordnet.
( M EE − 1) / τ E
M IE / τ I
M EI / τ E
( M II − 1) / τ I
(18)
Die Stabilität des Fixpunktes ist bestimmt durch die Realteile der Eigenwerte dieser
Matrix. Die Eigenwerte berechnen sich nach
M − 1 M II − 1
λ = EE
+
±
τI
τE
2
M EE − 1 M II − 1
4 M EI M IE
+
−
τI
τ Eτ I
τE
.
(19)
Wenn der Realteil beider Eigenwerte kleiner bzw. größer Null ist, dann ist der
Fixpunkt stabil bzw. instabil. Wenn der Ausdruck unter der Wurzel positiv ist, so sind
die Eigenwerte reell und das Verhalten Nahe dem Fixpunkt ist exponentiell. D.h. es
findet eine exponentielle Bewegung zum Fixpunkt hin statt, wenn beide Eigenwerte
negativ sind und vom Fixpunkt weg, wenn beide Eigenwerte positiv sind. Im Falle des
negativen Radikanden ist die Quadratwurzel eine imaginäre Zahl und die Eigenwerte
bilden ein komplex konjugiertes Paar. In diesem Falle ist das Verhalten des Systems
in der Nähe des Fixpunktes oszillierend und die Trajektorie windet sich bei negativem
Realteil der Eigenwerte spiralförmig zum Fixpunkt hinein oder bei positiven aus dem
Fixpunkt heraus. Der Imaginärteil des Eigenwertes bestimmt die Frequenz der
Oszillationen Nahe dem Fixpunkt. Der Real- und Imaginärteil von einem der
Eigenwerte ist in Abbildung 2B aufgezeichnet. Diese Darstellung zeigt, dass der
Fixpunkt bei τ I < 40ms stabil und bei größeren Werten für τ instabil ist.
Abbildung 3: Aktivität eines exzitatorisch-inhibitorischen Feuerratenmodells bei stabilem Fixpunkt.
(A) Abklingen der Frequenzschwankungen von
Phasenebenendarstellung.
v E und vI über die Zeit und (B) die zugehörige
10
Abbildung 3 und 4 zeigen Beispiele in denen der Fixpunkt stabil bzw. instabil ist. In
Abbildung 3A sind die Oszillationen von v E und vI gedämpft und die Feuerraten
bewegt sich zu einem stabilen Fixpunkt. Die zugehörige Phasenebenentrajektorie
stellt eine kollabierende Spirale dar ( Abbildung 3B ). In Abbildung 4A wächst die
Oszillation an und in Abbildung 4B ist die Trajektorie eine sich nach außen bis zu
einem Begrenzungskreis immer weiter ausdehnende Spirale. Ein Begrenzungskreis
ist eine geschlossene Umlaufbahn in der Phasenebene, die ein periodisches
Verhalten zeigt. Ohne Rektifizierung der Aktivierungsfunktionen würde sich die
Trajektorie spiralförmig bis ins unendliche ausdehnen. Die Nichtlinearität der
Rektifizierung verhindert, dass die Trajektorie über die Null hinaus expandiert und
stabilisiert auf diese weise den Begrenzungskreis.
Abbildung 4: Aktivität eines exzitatorisch-inhibitorischen Feuerratenmodells bei instabilem Fixpunkt.
(A) periodische Schwankungen der Feuerraten von
Phasenebenendarstellung.
v E und vI über die Zeit und (B) die zugehörige
Es gibt eine Vielzahl von Möglichkeiten wie ein nichtlineares System von einem
stabilen Fixpunkt zu einem Begrenzungskreis übergeht. Solche Übergänge werden
als Bifurkationen (Gabelungen) bezeichnet. Der zwischen Abbildung 3 und Abbildung
4 zu sehende Übergang ist eine Hopfbifurkation. In diesem Fall wird der Fixpunkt bei
Änderung eines Parameters (hier τ I ) instabil, wenn der Realteil des komplexen
Eigenwerts sein Vorzeichen ändert. In einer Hopfbifurkation entsteht der
Begrenzungskreis bei einer endlichen Frequenz, vergleichbar mit dem Verhalten
eines Typ II-Neurons, der anfängt Aktionspotentiale zu feuern. Andere
Bifurkationsformen wiederum zeigen ein Typ I-Verhalten mit Oszillationen bei der
Frequenz von Null. Ein Beispiel für diese Formen ist eine Sattelpunktbifurkation, die
dann entsteht, wenn sich die Parameter derart ändern, dass zwei Fixpunkte, einer
stabil und der andere instabil, sich im gleichen Punkt in der Phasenebene treffen.
Der Bulbus olfaktorius
Der Bulbus olfaktorius und analoge olfaktorische Regionen bei Insekten zeigen
Beispiele sensorischer Verarbeitung, die oszillatorische Aktivität zur Folge haben.
Der Bulbus olfaktorius ist die erste Stufe der Verarbeitung nach der
Geruchsrezeptorebene des Geruchssystems der Vertebraten. Die
Geruchsrezeptoren tragenden Neurone reagieren auf Duftstoffe und senden die
Information über ihre Axone zum Bulbus olfaktorius. Diese Axone enden in den
11
Mitral-, Pinselzellen und lokalen Interneuronen, die Knötchen bilden. Die Mitral- und
Pinselzellen übermitteln die Informationen zur p rimären olfaktorischen Hirnrinde und
haben synaptische Verbindungen zu den Körnerzellen, der größeren Gruppe
inhibierender Zellen. Die Körnerzellen wiederum inhibieren die Mitral- und
Pinselzellen.
Abbildung 5: (A) Aufzeichnung des extrazellulären Feldpotentials im Bulbus olfaktorius bei drei
aufeinander folgenden Riechvorgängen. (B) schematische Darstellung des Bulbus olfaktorius.
Bei vielen Vertebraten ist die Aktivität im Bulbus olfaktorius abhängig vom
Riechvorgang. Durch kurze Riechvorgänge gelangen die Duftstoffe zu den
Geruchsrezeptoren. Während drei kurzen aufeinander folgenden Riechvorgängen
sieht man in der Aufzeichnung des extrazellulär gemessenen Potentials drei
entsprechende große Ausschläge ( Abbildung 5). Uns interessieren hier jedoch die
kleineren Schwankungen höherer Frequenz, die durch oszillierende neurale
Aktivitäten entstehen und jeden der großen Ausschläge umgeben. Einzelne
Körnerzellen haben niedrige Feuerraten und feuern nicht bei jedem Zyklus der
Oszillation. In die phasensynchronen Oszillationen im Bulbus feuern verschiedene
Neurone zwar zu festen Zeitpunkten sind jedoch phasenverschoben zueinander.
Weiterhin werden durch verschiedenartige Düfte Unterschiede in den Amplituden und
Phasen hervorgerufen.
Li und Hopfield haben 1989 die Mitral- und Körnerzellen des Bulbus olfaktorius als
einen nichtlinearen inputgesteuerten Netzwerkoszillator modelliert. Abbildung 5B
zeigt die Architektur dieses Modells, dass die Gleichungen 14 und 15 (mit
M EE = M II = 0 ) benutzt . Diese fehlenden Verbindungen stimmen mit der Anatomie
des Bulbus überein und die Feuerraten von v E bzw. vI entsprechen denen von den
Mitral- bzw. Körnerzellen.
Die Abbildung 6A zeigt die Aktivierungsfunktionen des Modells. Die Zeitkonstanten
der Populationen von Körner- bzw. Mitralzellen sind gleich: τ E = τ I = 6.7 ms. h E
repräsentiert den von den Geruchsrezeptoren zu den Mitralzellen kommenden und
hI den konstanten vom olfaktorischen Kortex zu den Körnerzellen absteigenden
Input.
12
Abbildung 6: Aktivierungsfunktionen und Eigenwerte des Modells des Bulbus olfaktorius. (A)
Aktivierungsfunktionen der Mitral- ( FE ) und Körnerzellen ( FI ). (B) Real- (links) und Imaginärteil des
Eigenwertes, dass bestimmt ob das Modell ein Fixpunkt- oder Oszillationsverhalten annimmt.
Das Feldpotential in Abbildung 6A zeigt die Oszillationen während jedes einzelnen
Riechvorgangs ohne die dazwischen liegenden Phasen. Um das Model diesem
Aktivitätsmuster anzupassen, muss h E einen Übergang zwischen Fixpunkt und
schwingender Aktivität induzieren. Vor einem Riechvorgang muss das Netzwerk
einen stabilen Fixpunkt mit niedriger Aktivität annehmen. Mit Ansteigen von h E durch
einen Riechvorgang muss dieser ausgeglichene Zustand instabil werden und zu
einer oszillierenden Aktivität führen. Die Stabilitätsanalyse des Fixpunktes und das
Einsetzen der Oszillation ist vergleichbar mit unserer vorherigen Stabilitätsanalyse
des Modells verknüpfter exzitatorischer und inhibitorischer Neurone. Die
vergleichbarkeit beruht auf den Eigenwerteigenschaften der linearen
Stabilitätsmatrix. In diesem Fall umfasst die Stabilitätsmatrix den Beitrag der am
Fixpunkt berechneten Ableitungen der Aktivierungsfunktionen. Damit der Fixpunkt
instabil wird, muss der Realteil von mindestens einem der in dieser Analyse
berechneten Eigenwerte größer als 1 werden. Damit Oszillationen entstehen, muss
der Imaginärteil von mindestens einem der destabilisierenden Eigenwerte ungleich
Null sein. Diese Anforderungen prägen der Verbindung zwischen den Mitral- und
Körnerzellen sowie den Inputs Nebenbedingungen auf.
Die Abbildung 6B zeigt die Änderung des Real- und Imaginärteils des releva nten
Eigenwertes λ während eines Riechvorgangs. Nach etwa 100 ms wird der Realteil
von λ größer als 1. Der zu diesem Zeitpunkt gemessene Imaginärteil zeigt, dass er
im Netzwerk etwa 40 Hz hervorruft. Nach etwa 300 ms des Riechvorgangs sinkt der
Realteil von λ unter 1 und infolgedessen hören die Oszillationen auf. Der Input h E
erhöht bei diesem Vorgang zum einen durch Änderung des Fixpunktortes auf der
Aktivierungsfunktion in Abbildung 6A den Eigenwert λ über 1 und zum anderen
destabilisiert es ein bestimmtes Neuron, das dann zu oszillieren anfängt. Das
endgültige Muster oszillatorischer Aktivität ist sowohl durch den Input h E als auch
die Rückkopplungen des Netzwerks bestimmt.
Die Abbildung 7 zeigt das Verhalten des Netzwerks während eines Riechvorgangs
mit zwei unterschiedlichen Düften, die durch zwei unterschiedliche h E -Werte
repräsentiert werden. Die oberen Zeilen zeigen die Aktivität von vier Mitral- und die
unteren von vier Körnerzellen an. Die Amplituden und die Phasen der Oszillationen
13
zusammen mit den Erkennungsmerkmalen der beteiligten Mitralzellen zeigen eine
Signatur der Identität der gerochenen Düfte.
Abbildung 7: Aktivitäten von vier Mitral- (oben) und vier Körnerzellen (unten) während eines
Riechvorgangs von zwei unterschiedlichen Duftstoffen.
Oszillator ische Verstärkung
Zum Schluss betrachten wir ein Beispiel für Netzwerkoszillationen, indem wir die
Verstärkung des Inputs in einem rückwärtsgekoppelten Netzwerk betrachten. Zwei
Faktoren regeln die Menge von selektiver Verstärkung , die in rückwärtsgekoppelten
Netzwerken existieren kann. Die wichtigste Forderung an die rückwärtsgekoppelten
Gewichte ist, dass das Netzwerk so stabil ist, dass die Aktivität nicht unbeschränkt
anwachsen kann. Eine andere mögliche Anforderung deutet sich durch die
Abbildung 8A und B an.
Abbildung 8: Selektive Verstärkung in einem rückwärtsgekoppelten Netzwerk mit Rektifizierung. (A)
Aufzeichnung von
h (θ ), mit θ als den bevorzugten Winkel. (B) Gleichgewichtszustand des Outputs
v (θ ) , mit θ als den bevorzugten Winkel.
In der Abbildung 8 zeigt der Output eine abgestimmte Antwort obwohl der Input zum
Netzwerk eine nur von θ abhängige konstante Funktion ist. Ein abgestimmter Output
ohne einen abgestimmten Input könnte als Speicherungsmechanismus benutzt
werden. Jedoch ist dieser Mechanismus problematisch, denn würde beispielsweise
dieser Mechanismus in dem primär sensorischen Areal auftreten, so würde er eine
andauernde Wahrnehmung hervorrufen obwohl der Reiz nicht mehr da wäre. Die
Vermeidung dieses Mechanismus im Netzwerk würde die rückwärtsgekoppelten
Gewichten sowie das Ausmaß der Verstärkung, das unterstützt werden könnte,
begrenzen.
14
Li und Dayan haben 1999 gezeigt, dass diese Einschränkung durch die Benutzung
der Dynamik von Netzwerken mit gekoppelten inhibitorischen und exzitatorischen
Neuronen wesentlich gelockert werden kann. Die Abbildung 9 zeigt ein Beispiel mit
stetiger Neuronenkennzeichnung, das auf eine stetige Version der Gleichung 14 und
15 beruht. Der Input im abgestimmten Fall ist h E (θ ) = 8 + 5 cos( 2θ ) ( Abbildung 9C) und
nicht abgestimmten Fall h E (θ ) = 8 im (Abbildung 9C). Rauschen mit einer
Standardabeichung von 0.4 stört diesen Input. Der Input ist über die Zeit konstant.
Sowohl auf konstanten als auch auf abgestimmten Input oszilliert das Netzwerk.
Abbildung 9A zeigt den Durchschnittswert der Oszillationsaktivitäten der
Netzwerkneurone über die Zeit als eine Funktion ihrer bevorzugten Winkel zum einen
mit Abstimmung des Inputs auf Rauschen (durchgezogene Linie) und zum anderen
ohne Abstimmung des Inputs auf Rauschen (gestrichelte Linie).
Abbildung 9: Selektive Verstärkung in einem exzitatorisch-inhibitorischem Netzwerk. (A) Über die
Zeit gemittelte Antwortaktivität des Netzwerks auf eine abgestimmten (durchgezogene Linie) und nicht
abgestimmten Input (gestrichelte Linie). ’x’ und ’o’ markieren die 0° bzw. -37° Punkte die in (B) und (C)
dargestellt sind. (B) und (C) zeigen die Aktivitäten der Neurone, die 0° bzw. -37° bevorzugenden. Die
auf Gleichung 14 und 15 beruhenden Parameter sind τ E = τ I = 10ms, hI = 0, M EI = −δ (θ − θ ' ) / ρ 0 ,
M EE = (1/ πρ0 )[5.9 + 7.8 cos( 2(θ − θ ' )) ]+ , M IE = 13.3 / πρ0 , M II = 0 .
Auf den abgestimmten Input antworten die Neuronen auf eine hochgradig
angepasste und verstärkende Art und Weise. Bis auf die Verstärkung ist die
durchschnittliche Antwort der Neuronen auf den nicht abgestimmten Input
unabhängig von θ . Die Abbildung 9B und 9C zeigen die Aktivität der einzelnen
Neurone mit θ = 0° (´o`) bzw. θ = −37° (´x`) über die Zeit für den abgestimmten bzw.
nicht abgestimmten Input. Das Netzwerk bildet keine andauernde Wahrnehmung,
weil der Output auf ein nicht abgestimmten Input ebenfalls nicht abgestimmt ist. Im
Gegensatz dazu produziert eine nicht-oszi llierende Form dieses Netzwerks mit
τ I = 0 ein gleichmäßig abgestimmte n Output als Antwort auf einen nicht
abgestimmten Input mit einer rückwärtsgekoppelten Gewichte derselben Stärke. Auf
diese Weise kann das oszillierende Netzwerk in einer Ordnung von hochgradig
selektiver Amplifikation arbeiten ohne falsch abgestimmte Aktivität zu produzieren.
15