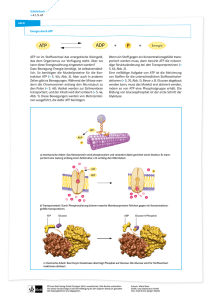
Glykolyse
Werbung

U. Albrecht BC1 Die Glycolyse (Glucose Katabolismus) 1. Übersicht über die Glykolyse 4. Kontrolle der Glykolyse 2. Reaktionschritte der Glykolyse 5. Stoffwechsel von anderen Hexosen 3. Gärung: Der anaerobe Weg des Pyruvats 6. Der Pentosephosphatweg U. Albrecht BC1 Gesamtübersicht Abbauweg von Glukose. Glykolyse (glykos = süss, lyse = auflösen) Glucose -> 2 Pyruvat Alternativweg für Glucoseabbau => Pentosephosphatweg U. Albrecht BC1 1. Übersicht Glykolyse Phosphorylgruppenübertragungsreaktionen Stufe I (Investition von Energie) -2 ATP Oxidationsmittel NAD+ + 2 NADH -> 2ATP NAD+ muss regeneiert werden Stufe II (Gewinnung von Energie) + 4 ATP U. Albrecht BC1 2. Die Reaktionschritte der Glykolyse A. Hexokinase: Verbrauch des ersten ATP Mg2+ schirmt neg. Ladungen ab -> γ-Phosphoratom wird zugänglich für nucleophilen Angriff. Glucose induziert Konformationsänderung der Hexokinase Kinase = Enzym das Phosphorylgruppen auf Metaboliten überträgt. Name des Metaboliten wird vorangestellt -> Hexokinase für Hexosen, Glucokinase in Leber um Blutglucosespiegel aufrecht zu halten. Magnesium essentiell !!! 2 Kieferbewegung -> ATP nähe C6 und Wasser weg -> keine Hydrolyse von ATP = substratinduzierte Konformationsänderung vrgl. Adenylat-Kinase U. Albrecht BC1 B. Glucosephosphat-Isomerase PGI = Phosphoglucose-Isomerase Aldose wird zu Ketose Ringöffnung und Ringschluss Säure-Base katalysiert durch das Enzym Schritt 1: Substrat bindet Schritt 2: Säure (ε-Aminogruppe von Lys) Schritt 3: Base (Carboxylatgruppe v. Glu) abstrahiert Proton Schritt 4: H+ an C1 => Protonenübertragung Schritt 5: Ringschluss U. Albrecht BC1 C. Phosphofructokinase: Verbrauch des zweiten ATP Ähnlich der Hexokinase Reaktion (Mg2+). PFK zentral in Glykolyse, da geschwindigkeitsBestimmender Schritt. AMP erhöht Aktivität ATP, Citrat allosterische Hemmer (siehe später). Bisphosphat und nicht diphosphat, da Phosphatgruppen getrennt. U. Albrecht BC1 D. Aldolase Spaltung von FBP (1xC6) in 2 C3 Einheiten Nummerierung der C Atome ändert = Aldolspaltung Mechanismus der basenkatalysierten Aldolspaltung Enolat-Zwischenprodukt entsteht -> Resonanzstabilisiert. G6P->F6P Isomerisierung da nur F6P gleiche in sich umwandelbare C3 Einheiten liefert die gleich abgebaut Werden können. 2 Klassen von Aldolasen Klasse I Aldolasen Klasse II Aldolasen U. Albrecht BC1 In Pilzen, Algen und Bakterien Keine Schiffsche Base sondern sondern zweiwertiges Kation Polarisiert Carbonylsauerstoff -> Stabilisierung des Enolats Klasse I Aldolasen in Tieren und Pflanzen Schritt 1: Substratbindung Schritt 2: Reaktion der Carbonylgruppe mit ε-Aminogruppe Bildung einer Schiffschen Base. Schritt 3: Spaltung der C3-C4 Bindung Bildung von Enamin Schritt 4: Protonierung des Enamins zu Iminiumkation Schritt 5: Hydrolyse, freies Enzym regeneriert U. Albrecht BC1 E. Triosephosphat-Isomerase (TIM) Base Säure Isomerisierung wie bei G6P->F6P Aldose->Ketose Übergangszustand bindet besser an Enzym als Substrat Aldose Ketose (Analoge zu Endiol, wie 2-Phosphoglykolat Binden besser an Enzym als Substrate) U. Albrecht BC1 Hefe TIM im Komplex mit 2-Phosphoglykolat flexible Schleife Glu α/β Fass. 8 β Faltblätter und 8 α Helices His Lys Stereoelektrische Kontrolle durch Flexible Schleife -> effektive Umsetzung des Substrates da Übergangszustand stabilisiert wird. Geschwindigkeit der Assoziation von Enzym und Substrat diffusionskontrolliert -> Bildung des Produkts so rasch wie Aufeinandertreffen von Enzym und Substrat -> GAP und DHAP immer im Gleichgewicht 2-Phosphogkykolat GAP weiter Umgesetzt -> Fliessgleichgewicht -> DHAP wird in GAP umGewandelt -> beide Produkte werden Im Stoffwechsel umgesetzt. U. Albrecht BC1 Übersicht 1. Stufe der Glykolyse F. Glycerinaldehyd-3-phosphat-dehydrogenase: Bildung des ersten energiereichen Zwischenprodukts U. Albrecht BC1 U. Albrecht BC1 Mechanistische Untersuchungen Cys-Sulfhydrylgruppe imAktiven Zentrum Direkte Hydridübertragung Acyl-Enzym Zwischenprodukt U. Albrecht BC1 Enzymmechanismus der GAPDH Schritt 1: GAP bindet an Enzym Schritt 2: Sulfhydrylgruppe greift Aldehydgruppe an und es entsteht ein Thiohalbacetal Schritt 3: Übertragung von Hydridion auf NAD+ -> Hemiacetal wird zu Ester oxidiert. Thioester hohe freie Enthalpie bei Hydrolyse -> Energie der Aldehydoxidation gespeichert in Form von NADH und Thioester. Schritt 4: NAD+ verdrängt NADH Schritt 5: Pi greift Thioester an -> Enzym wird regeneriert und 1,3 BPG entsteht. U. Albrecht BC1 G. Phosphoglycerat-Kinase: Produktion des ersten ATP Phosphoglycerat-Kinase PGK) 2 lappig ähnlich wie Hexokinase -> zusamenklappen -> wasserfreie Umgebung. Hexokinase und PGK Verschiedene Aminosäuresequenzen. 3PG U. Albrecht BC1 Verknüpfung der GAPDH- mit der PGK-Reaktion GAP + Pi + NAD+ ----> 1,3 BPG + NADH ∆G°‘ = +6.7 kJ/mol 1,3 BPG + ADP ----> 3PG + ATP ∆G°‘ = -18.8 kJ/mol _________________________________________________________________ GAP + Pi + NAD+ + ADP ----> 3PG + NADH + ATP ∆G°‘ = -12.1 kJ/mol Substratkettenphosphorylierung -> ATP Synthese ohne Sauerstoffbeteiligung Das NADH und O2 liefert durch die oxidative Phosphorylierung zusätzlich ATP U. Albrecht BC1 H. Phosphoglycerate-Mutase (PGM) Mutasen katalysieren intramolekulare Übertragungen einer funktionellen Gruppe von einer Position auf eine andere. Ist nicht direkt sondern über Phospho-His-Rest Energetisch neutrale Reaktion. U. Albrecht BC1 Postulierter Reaktionsmechanismus der Phosphoglycerat-Mutase Schritt 1: 3PG bindet an Enzym His ist phosphoryliert Schritt 2: Phosphorylgruppe wird übertragen Schritt 3&4: Komplex zerfällt U. Albrecht BC1 I. Enolase: Bildung des zweiten energiereichen Zwischenprodukts Dehydratisierung Mg2+ für Substratbindung an Enolase nötig. Fluorid ion (F-) blockiert Enolase und damit die Glykolyse. 2PG und 3PG reichern sich an. U. Albrecht BC1 J. Pyruvat-Kinase: Produktion des zweiten ATP Schritt 1: Phosphorylsauerstoff des ADP greift PEP-Phosphorylatom an -> ATP wird gebildet. Schritt 2: Tautomerisierung zu Pyruvate. U. Albrecht BC1 Als 2 Stufenprozess betrachtet -> Tautomerisierung setzt mehr Energie frei als PhosphorylgruppenÜbertragung -> Tautomerisierung treibt Reaktion. Zusammenfassung von Stufe II der Glykolyse U. Albrecht BC1 In Stufe 1 2 ATP investiert 4 ATP werden in Stufe 2 gewonnen (2x2 aus 2 GAP) 2 NADH gewonnen -> oxidative Phosphorylierung. 2 Pyruvat -> Citrat Zykuls (aerobe Bedingungen) U. Albrecht BC1 3. Gärung: Der anaerobe Weg des Pyruvats Muskel Milchsäuregärung Aerobe Bedingungen Hefe Alkoholische Gärung Anaerobe Bedingungen U. Albrecht BC1 A. Milchsäuregärung His 195 und Arg 171 orientieren Pyruvat im aktiven Zentrum Überschüssiges Lactat -> Umwandlung in Pyruvat oder Transport aus Muskel zu Leber-> Umwandlung von Lactat in Glucose Muskelkater nicht wegen Lactat Anhäufung sondern wegen Säureproduktion der Glykolyse Im Muskel bei hoher Aktivität und knapper Sauerstoffversorgung. LDH vollständig reversible Reaktion -> Gleichgewicht von Pyruvat und Lactat U. Albrecht BC1 B. Alkoholische Gärung Reaktion läuft bei der Herstellung von Bier und Wein ab. Auch bei Brot -> CO2 lässt Teig aufgehen. Thiaminpyrophosphat (TPP) essentieller Cofaktor der Pyruvatdecarboxylase (dieses Enzym in Tieren nicht vorhanden). Kann Carbanion stabilisieren wenn Proton abgegeben wird -> dipolares Carbanion oder Ylid U. Albrecht BC1 Schritt 1: Ylid C- greift Pyruvat an Schritt 2: Austritt von CO2 Bildung eines Carbanions Schritt 3: Protonierung Carbanion Schritt 4: Eliminierung Ylid Bildung von Acetaldehyd U. Albrecht BC1 Reduktion von Acetaldehyd und Regeneration von NAD+ Hefe (Yeast) ADH (=YADH) ist ein tetramer Jede Untereinheit bindet ein Zink-ion -> Polarisierung der Carbonylgruppe des Acetaldehyds -> stabilisiert Übergangszustand. Säugetiere haben ADH in der Leber = LADH Beseitigt baktereill produzierten Ethanol (Darmflora) Und von aussen eingenommenen Ethanol. -> Reaktion verläuft in umgekehrter Richtung. LADH ist ein Dimer wobei jedes Monomer 2 Zink Ionen bindet. U. Albrecht BC1 Energiebilanz der Gärung Pro Glucose: Gärung liefert 2 Moleküle ATP Geschwindigkeit: 100 x <-----> oxidative Phosphorylierung 38 ATP 1x Hefe unter anaeroben Bedingungen -> verbraucht mehr Glucose für Wachstum (= Pasteur Effekt). Muskeln: 2 typen von Fasern Typ I: schnelle Fasern, fast keine Mitochondrien -> wenig oxidative Phosphorylierung -> schnelle anaerobe Glykolyse. Typ II: langsame Fasern. Viele Mitochondrien. Oxidative Phosphorylierung. 4. Kontrolle der Glykolyse U. Albrecht BC1 A. Phospohfructokinase: Kontrolle der Glykolyse im Muskel Viele Reaktionen der Glykolyse verlaufen am Gleichgewicht -> Fliessgleichgewicht lässt Glykolyse Kontinuierlich laufen. Abrupte Änderung des Energiebedarfes -> Kontrolle 3 Reaktionen arbeiten fernab des Gleichgewichtes: Hexokinase, Phosphofructokinase (siehe slide 6) Pyruvat-Kinase Mg++ F6P T ATP Negative Änderung der freien Enthalpie -> Kandidaten für Kontrolle. Wenn Glycogen statt Glucose im Skelettmuskel verBraucht wird -> Hexokinase Reaktion umgangen R R/T Pyruvat-Kinase ist letzter Reaktionschritt in Glycolyse-> Nicht geeignet als Kontrollpunkt. Phospohfructokinase (PFK) geeignetster Kontrollpunkt 2 Untereinheiten Inhibitorstelle, Aktivatorstelle PFK = tetramer, R und T Konformationszustände ATP = Substrat und allost. Hemmstoff (hohe [ATP ]) AMP, ADP, F2,6P = Aktivatoren F6P = Substrat (R) U. Albrecht BC1 U. Albrecht BC1 Allosterische Änderung der PFK (Bacillus stearothermophilus) Aktivator R-Zustand T-Zustand Ionische Wechselwirkung -/+ F6P gebunden -/- F6P abgestossen Inhibitor (PGC= 2 Phosphoglykolat = PEP Analog) ATP wird durch Creatinkinase und Adenylat-Kinase Gepuffert. D.h. ATP Konzentrationsveränderungen Sind relativ gering -> d.h. kann nicht alleine die Aktivität von PFK regulieren. -> AMP und ADP heben Hemmung der PFK durch ATP auf da sie bevorzugt an den R-Zustand binden. U. Albrecht BC1 B. Substratkreislauf Änderung des Flusses in der Glykolyse 100 fach. Solche drastischen Steigerungen nur möglich wenn Reaktionen nahe an Gleichgewicht. Dies nicht der Fall für PFK -> Zweites Enzym das Rückreaktion katalysiert -> Fructose-1,6-Bisphosphatase (FBPase). PFK: F6P + ATP --> FBP + ADP -25.9 kJ/mol FBPase: FBP + H2O --> F6P + Pi -8.6 kJ/mol Kombination --> netto 1 ATP verbraucht Gruppe von gegenläufigen Reaktionen = Substratkreislauf oder Substratcyclus Aktivatoren der PFK (AMP, F2.6P) hemmen die FBPase. --> Aktivierung PFK --> Inhibierung FBPase Essentiell für plötzliche hohe glycolytische Flussrate bei hohem Energiebedarf (Fluchtbereitschaft usw.) Möglicherweise auch hormonell kontrolliert. Thermogenese (Schilddrüsenhormone). Chronische Obesitas -> geringerer Stoffwechselumsatz -> verringerte Thermogenese -> Kälteempfindlich (keine Erhöhung der Schilddrüsenhormone bei Kälte). Ruhezustand Muskel, beide Enzyme aktiv -> Fluss gering Aktiver Muskel -> PFK nimmt zu FBPase ab --> dramatischer Anstieg des glykolytischen Flusses U. Albrecht BC1 5. Stoffwechsel von anderen Hexosen als Glucose U. Albrecht BC1 A. Fructose Fructose Brennstoffquelle wenn Früchte oder Saccharose aufgenommen werden. Muskel und Leber verschiedene Repertoire an Enzymen -> Fructose Abbau verschieden. Zuviel Fructose (Infusion) -> F1P schneller produziert als abgebaut-> ATP Konentration in Leber sinkt -> Glykolyse und Lactatproduktion steigt -> hohes Lactat im Blut lebensbedrohlich. Fructoseunverträglichkeit -> Typ-B-Aldolase Mangel -> Individuen entwickeln Abneigung Gegenüber süssem. Kann für Synthese von Triacylglyceriden gebraucht Werden. U. Albrecht BC1 B. Galactose Milchprodukte -> Lactose = Disaccharid aus Glucose und Galactose. Hexokinase erkennt Galactose nicht -> Epimerisierungschritt Epimerisierung läuft über UDP-Glucose (siehe später). Galactoseabbau U. Albrecht BC1 Galactosämie Mangel an Galactose-1-phosphat uridyltransferase -> Anhäufing toxischer Nebenprodukte -> EntwicklungsStörungen, geistige Retardierung. Glalactose hoch im Blut -> in Augenlinse umgewandelt In Galactitol -> grauer Star Galacotsefreie Diät behebt alle Symptome. Für Synthese von Glycoproteinen Galactose aus Umkehrung der Epimerisierungsreaktion aus Glucose synthetisiert. U. Albrecht BC1 C. Mannose C2 Epimer von Glucose Hexokinase erkennt Mannose -> Phosphorylierung 6. Der Pentosephosphatweg U. Albrecht BC1 Stufe 1: oxidative Reaktionen Stufe 2: Isomerisierung und Epimeris. Stufe 3: Bilden und Spalten von C-C Stufen 2+3 vollständig reversibel Energiewährungen in der Zelle: ATP NADH -> für ATP Synthese NADPH -> reduktive Biosynthesen NAD+/NADH = 1000 Metabolitenoxidation NADP+/NADPH = 0.01 reduktive Biosynth. NADPH durch oxidation von G6P erzeugt, aber nicht in Glycolyse sondern in Separatem Weg -> Pentosephosphatweg Vor allem in Geweben mit Lipidbiosynthese: Leber, Milchdrüsen, Fettgewebe, Nebennierenrinde 30% der Glucoseoxidation in der Leber Über Pentosephosphatweg U. Albrecht BC1 A. Stufe 1: Oxidation unter Bildung von NADPH und Ribulose-5-phosphat Startpunkt G6P aus Glykolyse (Hexokinase) oder Glykogenabbau 1. Glucose-6-phosphate dehydrogenase -> Lacton 2. 6-Phosphogluconolactonase -> Ringöffnung U. Albrecht BC1 B. Stufe 2: Isomerisierung und Epimerisierung von Ribulose-5-phosphat Relative Mengen von R5P und Xu5P hängt vom Bedarf der Zelle ab. Wenn Pentosephosphatweg nur für NADPH Produktion gebraucht wird ist das Verhältnis Xu5P : R5P = 2:1 In teilenden Zellen DNA-Synthese gesteigert-> R5P vermehrt produziert da Vorläufermolekül für Nucleotide. U. Albrecht BC1 C. Stufe 3: Reaktionen zur Knüpfung und Spaltung von C-C Bindungen Aus 3 G6P entstehen 3 Ru5P. Aus diesen 3 C5 Zuckern entstehen dann 2 C6 und ein C3 Zucker. Transaldolasen und Transketolasen -> Bildung stabilisierter Carbanionen Mit Hilfe von TPP Transketolase Reaktion Katalysiert die Übertragung von C2-Einheiten U. Albrecht BC1 Transaldolase Reaktion Katalysiert die Übertragung von C3-Einheiten U. Albrecht BC1 Transketolase Transaldolase Transketolase U. Albrecht BC1 D. Kontrollmechanismen des Pentosephosphatweges Hauptprodukte des Pentosephosphatweges Glucose-6-phosphatdehydrogenase bestimmt Fluss durch Pentosephosphatweg. Dieser Schritt wird durch NADPH Konzentration kontrolliert.