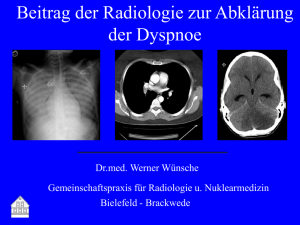
Tierärztliche Hochschule Hannover Retrospektive - Ti
Werbung

Tierärztliche Hochschule Hannover Retrospektive Untersuchungen zu Lungenveränderungen bei Weißbüschelaffen (Callithrix jacchus) in einer Versuchstierkolonie INAUGURAL – DISSERTATION zur Erlangung des Grades eines DOKTORS DER VETERINÄRMEDIZIN - Doctor medicinae veterinariae (Dr. med. vet.) vorgelegt von Marius Fabian Kunze Gelsenkirchen Hannover 2012 Wissenschaftliche Betreuung: Univ.-Prof. Dr. F.-J. Kaup Deutsches Primatenzentrum Göttingen, Abteilung Infektionspathologie 1. Gutachter: Univ.-Prof. Dr. F.-J. Kaup Tierärztliche Hochschule Hannover, Deutsches Primatenzentrum Göttingen 2. Gutachter: Univ.-Prof. Dr. M. Hewicker-Trautwein Tierärztliche Hochschule Hannover Institut für Pathologie Tag der mündlichen Prüfung: 16.11.2012 Der Übergang vom Affen zum Menschen sind wir. Konrad Lorenz 1 Einleitung 1 2 Literaturübersicht 2 2.1 Weißbüschelaffen 2 2.1.1 Systematik 2 2.1.2 Vorkommen und Lebensweise 2 2.1.3 Haltung in der Obhut des Menschen 3 2.2 Überblick über morphologische Lungenveränderungen 5 2.2.1 Hypostase 5 2.2.2 Atelektase 5 2.2.3 Emphysem 6 2.2.4 Kreislaufstörungen 7 2.2.5 Pigmentablagerungen und Stoffwechselstörungen 9 2.2.6 Entzündungen 11 2.2.6.1 Alveoläre Herdpneumonien 11 2.2.6.2 Interstitielle Pneumonien 14 2.2.6.3 Sonderformen 16 2.2.6.4 Granulomatöse Pneumonien 17 2.2.7 Tumoren 17 2.3 Respiratorische Pathogene bei nicht humanen Primaten 18 2.3.1 Viren 18 2.3.2 Bakterien 22 2.3.3 Parasiten 25 2.3.4 Pilze 27 3 Eigene Untersuchungen 30 3.1 Material und Methoden 30 3.1.1 Allgemeine Daten 30 3.1.2 Sektion und Probenentnahme 30 3.1.3 Histologische Aufarbeitung 31 3.1.4 Mikrobiologische und parasitologische Untersuchungen 32 3.1.5 Molekularbiologische Untersuchungen 32 3.1.5.1 RNA-Isolierung und Bestimmung der optischen Dichte 33 I 3.1.5.2 Reverse Transkription 34 3.1.5.3 Quantitative Echtzeit-PCR (qRT-PCR) 34 3.1.5.4 Nested PCR 37 3.1.6 Datenerfassung und -auswertung 39 3.1.7 Statistik 42 4 Ergebnisse 43 4.1 Allgemeine Angaben zu den untersuchten Weißbüschelaffen des DPZ 4.2 43 Überblick über die Lungenveränderungen im Tierbestand des DPZ 45 4.2.1 Untersuchung der minimalen Infiltrationen mit Entzündungszellen 53 4.2.2 Untersuchung der interstitiellen Pneumonien 57 4.3 Überblick über die Lungenveränderungen externer Tiere 68 5 Diskussion 72 6 Zusammenfassung 86 7 Summary 89 8 Literaturverzeichnis 92 9 Anhang 106 II ABKÜRZUNGSVERZEICHNIS Abb. Abbildung Aqua dest. destilliertes Wasser bzw. beziehungsweise c-DNA komplementäre DNA cm Zentimeter COPD chronic obstructive pulmonary obstruktive Lungenerkrankung) dNTP´s Desoxyribonukleosidtriphosphate DPZ Deutsches Primatenzentrum et al. und Mitarbeiter Fa. Firma g Gramm GALT gut associated lymphoid tissue (Darmschleimhautimmunsystem) GC-Gehalt Guanin- und Cytosingehalt HCl Chlorwasserstoff HE Hämatoxylin-Eosin i. m. intramuskulär KGW Körpergewicht KOH Kaliumhydroxid mg/kg Milligramm/Kilogramm min Minute mm Millimeter mM Millimolar ml Milliliter m 2 Quadratmeter m3 Kubikmeter ng Nanogramm nm Nanometer III disease (Chronische nmol Nanomol o. b. B. ohne besonderen Befund pmol Pikomol sec Sekunde sp. Spezies ssp. Subspezies Std. Stunde Tab. Tabelle z. B. zum Beispiel µl Mikroliter µm Mikrometer °C Grad Celsius IV Einleitung 1 Einleitung Der zur Familie der Krallenaffen gehörende Weißbüschelaffe (DICK u. DICK 1974; JENSON et al. 2002) zählt zu den Neuweltaffen und ist im ostbrasilianischen Regenwald beheimatet. Seine geringe Körpergröße, die im Vergleich zum Altweltaffen einfache Haltung, seine hohe Reproduktionsrate, geringe Haltungskosten und ein relativ geringes zoonotisches Risikopotenzial machen ihn zu einem häufig genutzten Tiermodell in der biomedizinischen Forschung. Im Rahmen einer Kooperation zwischen dem Fraunhofer Institut für Toxikologie und experimentelle Medizin (ITEM), Hannover und dem Deutschen Primatenzentrum (DPZ), Göttingen existiert ein Projekt, in dem der Weißbüschelaffe als Tiermodell für chronische Atemwegserkrankungen des Menschen, u. a. COPD und Asthma, etabliert wird. In diesem Zusammenhang werden morphologische Untersuchungen und pathophysiologische Studien im Respirationstrakt des Weißbüschelaffen durchgeführt. Ziel ist es, das Spektrum der spontanen Lungenveränderungen bei dieser Affenart zu definieren und pathologische Prozesse von möglichen „background lesions“ abzugrenzen. Ein weiterer Untersuchungsschwerpunkt liegt in der Bestimmung gesundheitsrelevanter Pathogene des Respirationstraktes beim Weißbüschelaffen, deren Kenntnis der Gewährleistung eines definierten Gesundheitsstatus der für Versuche verwendeten Tiere dient. In dieser Arbeit wird das Spektrum von Lungenveränderungen mit Schwerpunkt auf entzündlichen Lungenveränderungen im Bestand der Weißbüschelaffenkolonie des DPZ retrospektiv untersucht. Anhand von aktuellen Sektionsfällen wird darüber hinaus versucht, Hinweise auf eine mögliche infektiöse Ätiologie der entsprechenden Läsionen zu erhalten, insbesondere im Hinblick auf eine Beteiligung bakterieller oder viraler Pathogene. Die Kenntnis relevanter Pneumonieerreger und deren Prävalenz in Weißbüschelaffenkolonien soll als Grundlage für die Konzeption einer definierten spezifisch pathogen-freien Weißbüschelaffenkolonie im DPZ dienen, die darüber hinaus auf andere Weißbüschelaffenhaltungen im Versuchstierbereich übertragen werden kann. 1 Literaturübersicht 2 Literaturübersicht 2.1 Weißbüschelaffen 2.1.1 Systematik Die Krallenaffen gehören zu den auf dem amerikanischen Kontinent endemischen Neuweltaffen. Ihr Name ist zurückzuführen auf die Ausbildung von Krallen an Fingern und Zehen mit Ausnahme der Großzehe, die einen Nagel besitzt. Die Familie der Krallenaffen (Callitrichidae) umfasst die Gattungen der Tamarine (Saguinus), der Löwenäffchen (Leontopithecus), Zwergseidenäffchen (Cebuella) der und Springtamarine der Marmosetten (Callimico), der (Callithrix). Der Weißbüschelaffe, Callithrix jacchus, wird der letztgenannten Gruppe zugeordnet (WOLTERS u. IMMELMANN 1988; NEUSSER et al. 2001) 2.1.2 Vorkommen und Lebensweise Der Weißbüschelaffe stammt ursprünglich aus den tropischen und subtropischen Regenwäldern Brasiliens. Dank seiner hohen Anpassungsfähigkeit an vom Menschen veränderte Lebensräume gehört er trotz fortschreitender Dezimierung seiner ursprünglichen Habitate zu den wenigen nicht vom Aussterben bedrohten Krallenaffenarten (WOLTERS u. IMMELMANN 1988). Als obligater Baumbewohner ist er vollkommen an das Leben im Wald angepasst, meidet den Erdboden und kann sich nur über zusammenhängende Regelwaldgebiete ausbreiten (WOLTERS u. IMMELMANN 1988). Das morphologische Kennzeichen des Weißbüschelaffen sind seine beweglichen, vom Kopf abstehenden, weißen Ohrbüschel. Das Tier hat eine durchschnittliche Kopf-Rumpf-Länge von 20-25 cm (WOLTERS u. IMMELMANN 1988). Zusammen mit der Schwanzlänge verdoppelt sich die Körpergröße und beträgt ca. 50-60 cm bei einem Gewicht von ca. 350-450 g (ABBOTT et al. 2003). In freier Wildbahn leben die Tiere in Gruppen, die nicht selten bis zu 13 Mitglieder umfassen. Nach ROWE et al. (1996) ist die Sozialstruktur variabel, und es finden sich Gruppen aus einem Weibchen und mehreren Männchen oder aus einem Männchen und mehreren Weibchen oder auch aus mehreren Männchen und mehreren Weibchen. In menschlicher Obhut bilden sich innerhalb einer Gruppe 2 Literaturübersicht stabile Sozialverbände aus, wenn die Zucht mit dem Elterntierpaar beginnt und man die Nachkommen in der Elternfamilie belässt. Das Elterntierpaar bildet dann die Spitze der Rangordnung (WOLTERS u. IMMELMANN 1988). Rangniedere Weibchen helfen zwar bei der Aufzucht von Nachkommen, in Gegenwart des ranghöchsten Weibchens bleibt jedoch dessen Ovulation aus, wodurch Inzucht vermieden wird (HAIG 1999). Experimentell konnte nachgewiesen werden, dass kurze Zeit nach dem Entfernen des ranghöchsten Weibchens bei dem nächsten rangniederen Tier ein regelmäßiger Zyklus einsetzt (WOLTERS u. IMMELMANN 1988). Weißbüschelaffen sind die meiste Zeit des Tages mit dem Anlegen von Nagelöchern in Baumrinden beschäftigt. Diese füllen sich mit Baumexsudat, das hochwertige Kohlenhydrate und wichtige Mineralstoffe liefert und erheblich zur Ernährung der Weißbüschelaffen beiträgt. Ihren Eiweißbedarf decken die Tiere über Insekten, ihrer zweiten wichtigen Nahrungsquelle, an die sie sich wegen ihres geringen Körperumfangs fast unbemerkt anschleichen können. Vervollständigt wird das Nahrungsspektrum durch vitaminreiche Früchte und Blüten sowie Nektar (WOLTERS u. IMMELMANN 1988). 2.1.3 Haltung in der Obhut des Menschen Die Nutzung des Weißbüschelaffen als Modell für die biomedizinische Forschung entwickelte sich in den 70-er Jahren. Heute gehört er in europäischen Forschungseinrichtungen zu dem am häufigsten verwendeten nicht humanen Primaten (ABBOTT et al. 2003). Aufgrund seiner verwandtschaftlichen Nähe zum Menschen ist er ein beliebtes Tiermodell in der virologischen Onkologie, Reproduktionsphysiologie, Transplantationsimmunologie, Pharmakologie, Endokrinologie, Toxikologie, Teratologie, Verhaltenskunde, Osteologie und wird außerdem als Versuchstier bei der Erforschung bakteriologischer und viraler Infektionskrankheiten und autoimmuner Erkrankungen, wie z. B. der Multiplen Sklerose, eingesetzt (DAVID et al. 2009). In der Infektionsforschung respiratorische Erkrankungen dient mit der Weißbüschelaffe viraler und als bakterieller Tiermodell Genese. für Neben Parainfluenza- und Adenoviren (SUTHERLAND et al. 1986; WEVERS et al. 2011) 3 Literaturübersicht sind experimentelle Infektionen mit Bordetellen, Streptokokken, Klebsiellen und Bacillus anthracis (BASKERVILLE et al. 1983; SCHILLER et al. 1989; LUDLAGE u. MANSFIELD 2003; LEVER et al. 2008) beschrieben. Kurze Geburtenabstände und zwei bis drei Nachkommen pro Wurf tragen dazu bei, dass Marmosetten und Tamarine zu den Primaten mit der höchsten Fruchtbarkeitsrate zählen (JAQUISH et al. 1991; TARDIF et al. 2003). Da die Reproduktionsrate aber im Wesentlichen von der Futterzusammensetzung und den Haltungsbedingungen abhängt, ist für eine erfolgreiche Zucht ein gutes Management unerlässlich. Weißbüschelaffen werden als exsudativor bzw. insektivor eingestuft, wobei neben Insekten und Früchten Baumexsudate von Gummi- und Kautschukbäumen bis zu 15 % ihrer Nahrung ausmachen (HARRISON u. TARDIF 1994; ROWE u. ROWE 1996; POWER u. MYERS 2009). Um in Gefangenschaft diesen besonderen diätetischen Ansprüchen gerecht zu werden, wird hier das von Akazien gewonnene Nahrungsergänzungsmittel Gummi arabicum eingesetzt, bei dem es sich um ein komplexes Polysaccharid handelt, das verschiedene Zucker und wichtige Mineralien enthält (BURROWS u. NASH 2010). Für optimale Haltungsbedingungen müssen einige grundsätzliche Voraussetzungen eingehalten werden. So ist es wegen des ausgeprägten Territorialverhaltens der Tiere nicht empfehlenswert, gleichgeschlechtliche, adulte Tiere zusammenzuhalten (FOX et al. 1984). Es empfiehlt sich vielmehr, Zuchtpaare einzeln in Käfigen unterzubringen (HEARN 1983). Da Weißbüschelaffen baumlebende Tiere sind und in vertikale Richtung flüchten, ist die Höhe des Geheges wichtiger als die Bodenfläche. So liegen nach den aktuellen Leitlinien der Europäischen Kommission für die Unterbringung und Pflege von Versuchstieren die Käfigmindesthöhe bei 1,5 m und die Mindestbodenfläche für 2 Tiere bei 0,5 m². Um dem natürlichen Verhaltensmuster der Tiere gerecht zu werden, sollte auf Ausgestaltungselemente im Haltungsbereich geachtet werden, wie z. B. Sitzstangen, Plattformen, Seile und Nestboxen. Das Temperaturoptimum liegt bei dieser tropischen Affenart zwischen 23 und 28 °C und die Luftfeuchtigkeitswerte zwischen 40 und 70 % (ETS 124 2007). 4 Literaturübersicht 2.2 Überblick über morphologische Lungenveränderungen Die im folgenden verwendete Klassifikation der Lungenerkrankungen erfolgte anhand der gängigen humanpathologischen Nomenklatur (BÖCKER 2008), modifiziert nach dem veterinärmedizinischen Lehrbuch „Pathologic Basis of Veterinary Diseases“ (MCGAVIN u. ZACHARY 2007). 2.2.1 Hypostase Bei der Hypostase handelt es sich um eine postmortale, passive Blutfülle der Lunge, welche meist schon in der Agonie einsetzt. Abhängig von der Seite, auf welcher das Tier nach dem Tod liegt, sammelt sich das Blut entweder in der linken oder der rechten Lungenhälfte. Histopathologisch stellen sich vor allem Venen, Venolen und Kapillaren dilatiert und stark blutgefüllt dar (SMITH et al. 1978; DAHME u. WEISS 2007). 2.2.2 Atelektase Bei einer Atelektase liegt eine unvollständige Ausdehnung der Lunge vor, bei der das Lungengewebe gar keinen oder einen wesentlich reduzierten Luftgehalt aufweist. Im Embryonal- und Fetalleben enthalten die Lungen keine Luft (totale fetale Atelektase). Dehnt sich das Lungengewebe nach der Geburt nicht aus, handelt es sich um eine angeborene Form der Atelektase. Diese ist von der erworbenen Form abzugrenzen, welche im Rahmen pathologischer Prozesse auftritt, und bei der lufthaltige Bereiche wieder luftleer sind. Bei der erworbenen Form wird entsprechend der Entstehungsursache eine Obstruktionsatelektase von einer Kompressionsatelektase unterschieden. Die Ursachen von Obstruktionsatelektasen sind Verlegungen der Luft leitenden Wege z. B. durch entzündliche Prozesse oder Aspiration von Fremdkörpern oder körpereigenen Stoffen. Die Ausprägung ist abhängig von der kollateralen Versorgung und dem Ausmaß der Obstruktion. Kompressionsatelektasen sind entweder bei raumfordernden Prozessen in der Pleurahöhle, wie Abszessen und Tumoren, oder bei Druckerhöhungen, wie durch Chylothorax, Hydrothorax oder Empyemen, zu erwarten. Makroskopisch erscheinen atelektatische Lungenabschnitte dunkelblau 5 Literaturübersicht und eingesunken. Histopathologisch stellen sich die Alveolen kollabiert mit parallel verlaufenden Alveolarwänden dar, wodurch das interstitielle Gewebe prominent erscheint (DAHME u. WEISS 2007; MATHIS 2007; CASWELL u. WILLIAMS 2008; MCGAVIN u. ZACHARY 2009). 2.2.3 Emphysem In der Humanmedizin bezeichnet das pulmonale Emphysem eine primäre Erkrankung, die als abnormale, permanente Vergrößerung der Luftwege distal der terminalen Bronchiolen beschrieben wird und mit Untergang alveolärer Strukturen durch Ruptur intraalveolärer Septen vergesellschaftet ist. In der tiermedizinischen Terminologie treten pulmonale Emphyseme als reversible oder irreversible Erweiterungen der Luft führenden Wege distal der terminalen Bronchiolen auf, welche nicht, wie in der Humanmedizin, primären Ursprungs sind, sondern immer sekundär infolge von Obstruktionen entstehen. Abhängig vom betroffenen Lungenbereich kann ein Emphysem als alveolär oder interstitiell klassifiziert werden. Alveoläre Emphyseme lassen sich entsprechend ihrer Lokalisation in herdförmige und diffuse Emphyseme unterteilen, wobei die akuten Emphyseme mit reversiblen und die chronischen Emphyseme mit irreversiblen Veränderungen einhergehen. Als Ursache für die akuten, herdförmigen Emphyseme kommen Stenosen und Verschlüsse von kleinen Bronchien oder Bronchiolen, z. B. durch Schleim oder entzündliche Exsudate, in Betracht (Obstruktionsemphyseme). Da der inspiratorische Druck den exspiratorischen übersteigt, sammelt sich Luft distal dieser Ventilstenose, woraus eine Überdehnung und Ruptur der Septen resultiert. Akuten, diffusen Emphysemen liegt ebenfalls eine Ventilstenose zugrunde, die z. B. mit einem anaphylaktischem Schock einhergeht. Chronische Lungenemphyseme werden bei chronischen Entzündungen und Ventilstenosen beschrieben und sind aufgrund des Parenchymschwunds irreversibel. Bei interstitiellen Emphysemen tritt Luft aus dem alveolären Bereich in das Lungenzwischengewebe und bildet unter der Pleura perlschnurartig angeordnete Bläschen. Mikroskopisch stellen sich die Alveolen, im Vergleich zum normalen Lungengewebe, stark erweitert dar mit dünn ausgezogen oder gerissenen Alveolarsepten, deren Reste als teils auffällige, stummelförmige 6 Literaturübersicht Fortsätze in die Hohlräume hineinreichen (SANDRITTER et al. 1977; DAHME u. WEISS 2007; CASWELL u. WILLIAMS 2008). 2.2.4 Kreislaufstörungen Hyperämie Aktive Hyperämien treten bei akuten Entzündungsprozessen auf, wohingegen passive Hyperämien auf einen gestörten Abfluss des Blutes aus der Lunge zurückzuführen sind. Abhängig vom Alter des Prozesses teilt sich die passive Stauungshyperämie in die akute und die chronischen Form auf. Zu akuten Verläufen kommt es bei Linksherzinsuffizienzen. Lungenstauungen besonders infolge einer Dagegen werden chronische lang anhaltenden Stenose oder Insuffizienz der Mitralklappe beschrieben. Makroskopisch sind Gewicht und Volumen der Lunge erhöht, ihre Farbe ist dunkel-blaurot und die Schnittfläche feucht und blutig. Histologisch lassen sich je nach Stadium stark gefüllte Kapillaren und hämosiderinreiche Makrophagen (Herzfehlerzellen) feststellen. Lungenindurationen und degenerative Gefäßwandveränderungen sind häufige Folgeerscheinungen (DAHME u. WEISS 2007; MCGAVIN u. ZACHARY 2007). Lungenödem Ein Lungenödem bezeichnet eine abnormale Ansammlung seröser Flüssigkeiten in den Alveolen und/oder im interstitiellen Lungengewebe (alveoläres und interstitielles Lungenödem). Dies geschieht, wenn die Rate der Flüssigkeitstranssudation aus den Lungengefäßen die Rate des lymphatischen und alveolären Abflusses übersteigt. Makroskopisch stellt sich das akute Lungenödem in Form einer rötlich bis blassen Verfärbung der Lunge dar, wobei die Lunge feucht und schwer erscheint und von der Schnittfläche, besonders bei Druckanwendung, feinschaumige Flüssigkeit abfließt. Im chronischen Stadium nimmt die Viskosität der Flüssigkeit durch desquamiertes Alveolarepithel zu und ist von trüber Farbe. Neben neurogenen, kardiogenen, entzündlichen oder toxischen Ursachen können Lungenödeme auch agonal auftreten (DAHME u. WEISS 2007; MCGAVIN u. ZACHARY 2009). 7 Literaturübersicht Hyaline Membranen Hyaline Membranen stellen eine besondere Form des Lungenödems dar, bei der histologisch nachweisbare Glukosaminoglukan-Protein-Kondensate die Alveolen und Bronchiolen auskleiden. In der Humanmedizin bei Neugeborenen als idiopathisches Atemnot-Syndrom bekannt, sind sie in der Veterinärmedizin unter anderem in Verbindung mit interstitiellen Pneumonien beschrieben. So treten hyaline Membranen bei Weißbüschelaffen z. B. bei experimentellen Coronavirusinfektionen auf, bei denen der Krallenaffe als Tiermodell für SARS-Studien dient. Hier mischen sich abgesonderte Plasmaproteine mit Lipiden und Surfactant-Bestandteilen und bilden lang gezogene Membranen, die sich teilweise an alveoläre und bronchioläre Wände anheften und sich histopathologisch als eosinophiles, homogenes, amorphes Material darstellen (GREENOUGH et al. 2005; DAHME u. WEISS 2007; MCGAVIN u. ZACHARY 2009). Blutungen Die verschiedenen Formen von Lungenblutungen werden nach ihren Ursachen in Rhexis-, Arrosions- sowie Diapedesisblutungen unterteilt. Rhexisblutungen entstehen durch Traumata, wie z. B. Rippenfrakturen oder Stichverletzungen. Arrosionsblutungen können gelegentlich bei Lungengangrän oder Thrombembolien beobachtet werden. Für Diapedesisblutungen sind verschiedene toxische, hypoxische, kreislaufbedingte oder infektiöse Ursachen verantwortlich, die die Gefäßwandintegrität stören (DAHME u. WEISS 2007). Lungenembolie Zu den Lungenembolien gehören unter anderem die Thrombembolien, septische Embolien, Fettembolien sowie Tumorzellembolien. Thrombembolien der Lunge beruhen im Allgemeinen auf einem Thrombus, der sich im venösen Kreislauf befindet und Fragmente ablöst, die sich im Kapillarbett der Lunge verfangen. Die Gewebsläsionen hängen in erster Linie von der Größe und Anzahl der Emboli ab. Kleine, sterile Thromben sind klinisch und pathologisch unbedeutend, da sie durch das fibrinolytische System abgebaut werden. Bei größeren Thromben sind die 8 Literaturübersicht Veränderungen durch Blutungen und Nekrose gekennzeichnet. Histopathologisch lassen sich neben einem Saum aus Entzündungszellen zahlreiche Siderophagen im nekrotischen Gewebe nachweisen. Bleibt der Thrombus steril, kann die Läsion unter Bildung von Narbengewebe abheilen. Wenn Thrombusteile mit Pilzen oder Bakterien kontaminiert sind, handelt es sich um septische Embolien, bei denen massive Lungenödeme, embolisch-eitrige Bronchopneumonien und ausgedehnte Lungenabszesse die Folge sind. Fettembolien treten nach Knochenfrakturen oder Quetschungen von subkutanem Fettgewebe auf. Fetttröpfchen gelangen dabei in die eröffneten Venen des Verletzungsgebietes und mit dem Blut in die Lungenkapillaren. Wenn sich Tumorzellen in größerem Umfang in der Lunge festsetzen, handelt es sich um eine krebsige Pneumonie. Solche Tumorzellembolien können die Lungenfunktion erheblich beeinträchtigen und eine Todesursache bei bösartigen neoplastischen Prozessen darstellen (DAHME u. WEISS 2007; MCGAVIN u. ZACHARY 2009). 2.2.5 Pigmentablagerungen und Stoffwechselstörungen Pigmente mit Relevanz für den Respirationstrakt werden in zwei Gruppen unterteilt. Während Kohlenstaub- und Formalinpigmente die Gruppe der exogenen Pigmente bilden, zählen Melanin, Hämosiderin, und parasitäres Hämatin zu den endogenen Pigmenten. Kohlenstaub kommt ubiquitär in der Luft vor, wird nach aerogener Aufnahme von Makrophagen phagozytiert und sammelt sich in den regionären tracheobronchialen Lymphknoten. Da elementarer Kohlenstaub nicht weiter verstoffwechselt wird, bleibt er als schwarzes Pigment lebenslang reaktionslos im Gewebe liegen und lässt sich histopathologisch extra- sowie intrazellulär in Makrophagen nachweisen. Makroskopisch können die Lungen feine, subpleurale schwarze Verfärbungen aufweisen, die besonders bei entbluteten Lungen sichtbar sind. Diese als Anthracosis pulmonum bezeichnete Pigmentablagerung wird bei Haustieren und beim Menschen beschrieben. Bei nicht humanen Primaten liegen vor allem Fallberichte für Altweltaffen wie Weißlid-Mangaben (Cercocebus torquatus atys), Indische Hutaffen (Macaca radiata), Rhesusaffen (Macaca mulatta) und Javaneraffen 9 Literaturübersicht (Macaca fascicularis) vor (CHOUDARY et al. 1986; SHIMOI et al. 1998; DAHME u. WEISS 2007; MCGAVIN u. ZACHARY 2009; SEEMA et al. 2010). Derartige Fälle sind bei Weißbüschelaffen nicht publiziert. Beim Formalinpigment handelt es sich um einen histologischen Artefakt. Dieses entsteht, wenn Hämoglobinabbauprodukte mit saurer Formalinlösung in Verbindung kommen, besonders dann, wenn es zwischen Todeszeitpunkt und Gewebsfixierung zu Verzögerungen kommt. Makroskopisch nicht sichtbar stellt sich dieses Pigment histopathologisch als braun bis schwarz, fein und körnig dar (MCGAVIN u. ZACHARY 2009). Eine ausgeprägte Melaninablagerung im Gewebe wird als Melanosis maculosa bezeichnet und ist in der Lunge bei verschiedenen Haussäugern sowie beim Menschen beschrieben. Bei dieser kongenitalen Melanose, die klinisch ohne Relevanz ist, finden sich multifokale Pigmentablagerungen subpleural und im Lungenparenchym. Während pulmonale Ausprägungen bei nicht humanen Primaten noch nicht beschrieben wurden, sind neurokutane Ansammlungen dieses Pigments bei Javaneraffen bekannt (CHEN et al. 2009; MCGAVIN u. ZACHARY 2009). Bei Lungenhämosiderosen handelt es sich um Ablagerungen von Eisen-ProteinKomplexen als Folge chronischer Lungenstauungen, Lungenblutungen oder Teilerscheinungen allgemeiner Hämosiderosen nach hämolytischen Anämien. Chronische Lungenstauungen stehen in Verbindung mit einer unzureichenden Blutzirkulation und einer daraus resultierenden passiven Hyperämie in der Lunge. Lysierte Erythrozyten werden in den Bereichen, wo sie aufgrund des verlangsamten Blutflusses ihr Lebensende erreicht haben, von Alveolarmakrophagen phagozytiert. In diesen auch als Herzfehlerzellen bezeichneten Makrophagen stellt sich histopathologisch das Hämosiderin als goldgelbes bis goldbraunes, intrazelluläres, feingranuläres Pigment dar (DAHME u. WEISS 2007; MCGAVIN u. ZACHARY 2009). Die beim Altweltaffen weitverbreitete Lungenmilbe (Pneumonyssus simicola) bildet, vermutlich aus verstoffwechseltem Hämoglobin, parasitäres Hämatin, das sich als gelb-braun-schwarzes, anisotropes Pigment im Gewebe ablagert und eine Entzündung aufrechterhält, selbst wenn lebende Milben in der Lunge nicht mehr nachgewiesen werden können. Infektionen mit Lungenmilben sind bei Altweltaffen wie Rhesusaffen, Javaneraffen und Rotschwanzmeerkatzen 10 (Cercopithecus Literaturübersicht ascanius) bekannt (ANDRADE u. MARCHEVSKY 2007; MCGAVIN u. ZACHARY 2009; KOORIYAMA et al. 2010; SHIRAI u. GEOLY 2010). Für Neuweltaffen liegen keine Fallberichte vor. Zu den Stoffwechselstörungen Lungengewebes. Metastatische zählen unter Verkalkungen anderem der Verkalkungen Lungen resultieren des aus hyperkalzämischen Zuständen und führen zu dem makroskopischen Bild der Bimssteinlunge, wobei das diffuse, mineralisierte Lungengewebe stark verdichtet ist und eine reibeisenartige Konsistenz besitzt. Histopathologisch gehen diese Veränderungen auf Verkalkungen der alveolären Basalmembranen zurück. Ursächlich für die Hyperkalzämie sind vier Gründe. Nierenversagen mit sekundärem, renalem Hyperparathyreoidismus, eine Vitamin-DHypervitaminose, eine Synthese von Parathormon oder Parathormon verwandtem Protein im Rahmen von neoplastischen Erkrankungen oder massiver Untergang von Knochen durch primäre oder metastatische Neoplasien. Dystrophische Verkalkungen entwickeln sich fokal nach Lungenparenchymnekrosen, wie z. B. bei Granulomen, Tuberkeln oder in der Umgebung von abgestorbenen Parasiten, bei denen untergehende Zellen den Einstrom von Kalzium nicht kontrollieren können, was in einer Akkumulation von Kalzium im Zytosol resultiert und über pH-WertAbsenkungen im umliegenden Gewebe zu Kalziumausfällungen führt (MCGAVIN u. ZACHARY 2009). Im Anschluss an entzündliche Prozesse oder Verkalkungen kann es infolge einer Metaplasie von Bindegewebszellen ebenfalls zu diffusen oder herdförmigen Lungenverknöcherungen kommen. Makroskopisch gleicht diese diffuse Verknöcherung dem Bild der Bimssteinlunge (DAHME u. WEISS 2007). 2.2.6 Entzündungen 2.2.6.1 Alveoläre Pneumonien Bei den alveolären Pneumonien erreicht das Pathogen die Lunge auf aerogenem Infektionsweg. Eine Schädigung kann in den bronchialen und bronchiolären Regionen auftreten, durch zentrifugale Erregerausbreitung ebenfalls auf die Alveolen übergreifen oder nur in diesen auftreten. Abhängig von der Lokalisation des Entzündungsprozesses entwickelt sich entweder eine Lobärpneumonie, häufig die 11 Literaturübersicht Alveolen des ganzen Lungenlappen betreffend, oder eine Bronchopneumonie, herdförmig im Bereich der Bronchiolen, wobei Mischformen ebenfalls auftreten können. Bei Mitbeteiligung der Pleura handelt es sich um eine Pleuropneumonie. Während beim Menschen Streptococcus pneumoniae den Haupterreger von Lobärpneumonien darstellt, sind bei Tieren vor allem Pasteurella multocida und Mykoplasmen für diese Pneumonieform verantwortlich. Als Ursache eitriger Bronchopneumonien in der Humanmedizin sind neben Staphylokokken und Pneumokokken ebenfalls Klebsiellen, Pseudomonaden und Mykoplasmen bekannt, wohingegen in der Veterinärmedizin eine Vielzahl von bakteriellen Pathogenen, u. a. Trueperella pyogenes, Escherichia (E.) coli, Bordetella bronchiseptica oder Salmonellen bei dieser Pneumonieform beschrieben wurden. Lobärpneumonien Das makroskopische Bild der Lobärpneumonie, die in der veterinärmedizinischen Nomenklatur als fibrinöse Pneumonie bezeichnet wird, hängt vom Alter und Schweregrad der Läsion ab. Schwere Stauungen und Hämorrhagien dominieren in der frühen Phase. Später beginnt sich fibrinöses Exsudat auf der Pleuraoberfläche abzusetzen, das der Pleura ein stumpfes Aussehen verleiht. Die Farbe des Exsudates variiert je nach dem Anteil von Erythrozyten, Hämoglobin und Leukozyten. Im Zeitablauf der Lobärpneumonie lassen sich die Stadien der Anschoppung (vergrößerter, schwerer, dunkelroter Lungenlappen mit deutlicher Hyperämie), der roten Hepatisation (neben Kapillarhyperämie dichtes Fibrinnetz in den Alveolen) und der grauen Hepatisation (mit zahlreichen Granulozyten, die das Fibrinnetz auflösen) unterscheiden. Durch ein Nebeneinander von normalem und entzündlich verändertem Gewebe verschiedenen Alters erhält die Lunge makroskopisch das für die Lobärpneumonie charakteristische bunte, marmorierte Aussehen. Histologisch folgt auf die im Anfangsstadium auftretende massive Exsudation von Plasmaproteinen mit Verlegung von Alveolen und Bronchiolen eine Einwanderung von neutrophilen Granulozyten und Makrophagen, die mit Hilfe von Fibrin das flüssige Exsudat ersetzen und eine Ausheilung ermöglichen. Abhängig vom Ausmaß der Läsion kann diese jedoch mit deutlicher Fibrose, Pleuraadhäsion und 12 Literaturübersicht obliterierten Bronchien einhergehen. Ursächlich kommen für diese Pneumonieform vor allem Pasteurella multocida und Mykoplasmen in Betracht (BERENDT et al. 1978; DAHME u. WEISS 2007; MCGAVIN u. ZACHARY 2009; ABEE et al. 2012). Bronchopneumonien Wie bei der Lobärpneumonie erfolgt die Infektion bei der Bronchopneumonie ebenfalls auf aerogenem Weg und führt zu Bronchitis und Bronchiolitis, wobei die Lunge vor allem durch eine kranioventrale Konsolidierung gekennzeichnet ist. Hierbei handelt es sich um bakterielle Infektionen (meist im Anschluss an eine primäre Virusinfektion) mit Staphylokokken, Streptokokken, Trueperella pyogenes, E. coli, Bordetella bronchiseptica oder Salmonellen. Makroskopisch zeichnen sich die veränderten Lungenbereiche durch multiple, unregelmäßig verteilte rötliche Herde aus, zwischen denen normales, atelektatisches oder emphysematöses Gewebe zu finden ist. Von der Schnittfläche, insbesondere aus den Bronchien, fließt auf Druck eitriges Exsudat ab, weshalb diese Pneumonieform auch als katarrhalisch eitrige Bronchopneumonie bezeichnet wird. Histopathologisch sind Bronchopneumonien durch neutrophile Granulozyten, Makrophagen und zellulären Debris gekennzeichnet, der sich vor allem in den Lumina von Bronchien, Bronchiolen und Alveolen befindet. Bei einer deutlichen Desquamation von Alveolarepithelien liegt eine Alveolarzell- oder Desquamativpneumonie vor. Im weiteren Verlauf der Entzündung können einzelne Herde konfluieren, so dass größere Bereiche betroffen sind. In diesem Fall spricht man von einer konfluierenden Bronchopneumonie (DAHME u. WEISS 2007; MCGAVIN u. ZACHARY 2009). Aspirationspneumonien Anders als in der Veterinärpathologie, wo die Aspirationspneumonie mit den embolisch-eitrigen Pneumonien eine eigene Sonderform der Lungenentzündungen bildet, zählt sie nach der verwendeten humanen Klassifikation zu den Bronchopneumonien. Eine Aspiration von Fremdstoffen (z. B. infolge fehlerhaften Einführens von Magensonden) oder körpereigenen Stoffen (eitrige Entzündungen im Bereich der 13 Literaturübersicht oberen Luftwege, Fruchtwasser, Mageninhalt) führt meist zu einer purulenten bis abszedierenden Bronchopneumonie der kranioventralen Lungenabschnitte. Schädigend auf das Lungengewebe wirkt sich neben der direkten Reizwirkung des aspirierten Materials ebenso die fermentative Wirkung der im Aspirat enthaltenen saprophytären Mischflora aus. Im Folgenden kann sich durch Gewebseinschmelzung aus der eitrig-nekrotisierenden Pneumonie ein Lungengangrän entwickeln, das auf die Pleura übergreift und in ein jauchiges Empyem übergeht (DAHME u. WEISS 2007). 2.2.6.2 Interstitielle Pneumonien Anders als bei den alveolären Herdpneumonien erreichen hier die Pathogene die Lungen nicht ausschließlich auf aerogenem Weg. Neben einer Inhalation toxischer Gase (Ozon, Stickstoffmonoxid) oder anderer toxischer Substanzen sowie pneumotrope- bzw. epitheliotrope Viren und Bakterien kommt ebenfalls eine hämatogene Schädigung des Gefäßendothels, z. B. bei Septikämien oder durch Toxine, in Betracht. Die Läsionen und die daraus resultierenden Entzündungsprozesse finden primär in einer der drei Schichten der Alveolarwände statt, wobei aerogene Toxine Typ 1- und Typ 2-Pneumozyten und hämatogene Toxine vor allem das alveoläre Kapillarendothel oder die alveoläre Basalmembran schädigen. Die akute Phase der interstitiellen Pneumonie ist gekennzeichnet durch eine Exsudation proteinreicher Flüssigkeit, die sich in dem Alveolarraum mit Surfactant-Bestandteilen vermischt und hyaline Membranen bilden kann. Die Infiltration mit Entzündungszellen führt zur charakteristischen Verdickung der Alveolarwände, die die interstitielle Pneumonie kennzeichnet. In der nun folgenden proliferativen Phase werden nekrotische Typ 1-Pneumozyten durch hyperplastische Typ 2-Pneumozyten ersetzt. Im subakuten Krankheitsstadium handelt es sich bei den vorherrschenden Entzündungszellen nicht mehr um polymorphkernige, sondern um mononukleäre Zellen. Die chronische Phase ist durch reparative Prozesse mit Nachweis von Fibrosen im Interstitium sowie durch Persistenz von hyperplastischen Typ 2-Pneumozyten gekennzeichnet. Abhängig vom Schweregrad können akute interstitielle Pneumonien nur mit geringgradigen respiratorischen Symptomen 14 Literaturübersicht einhergehen und nach Abheilung keine signifikanten Folgeläsionen hinterlassen, wodurch sich ihre Diagnose in der Sektion als schwierig gestaltet. Häufig überdecken bakterielle Sekundärinfektionen das ursprüngliche Bild der interstitiellen Pneumonie. In diesen Fällen kann histologisch in den makroskopisch unverändert erscheinenden Lungenbereichen der primär interstitielle Entzündungscharakter noch nachgewiesen werden (DAHME u. WEISS 2007; MCGAVIN u. ZACHARY 2009). In der Humanpathologie werden, ähnlich wie in der Veterinärpathologie, die interstitiellen Pneumonien ihrer Ätiologie entsprechend in infektiöse, medikamentöse, metabolische, zirkulatorische, physikalische und immunologische Ursachen unterteilt. Keine Anwendung in der Veterinärpathologie findet die von der European Respiratory Society und American Thoracic Society (ERS/ATS) erstellte Klassifikation der idiopathischen chronischen interstitiellen Pneumonien. Diese Gruppe der fibrotischen Lungenerkrankungen wird im Humanbereich anhand pathomorphologischer und klinischer Untersuchungsmethoden sowie bildgebender Verfahren in sechs weitere Kategorien unterteilt, ohne dass hier näher darauf eingegangen werden soll (COSTABEL 2000; DACIC u. YOUSEM 2003; BÖCKER 2008). Eine Übersicht über Viren und Bakterien, welche unter anderem an interstitiellen Pneumonien beteiligt sein können, zeigt Tab. 1. 15 Literaturübersicht Tab. 1: Erregerbeispiele für die verschiedenen Pneumonieformen bei Haustieren und bei nicht humanen Primaten. Pneumonietyp Erregerbeispiel Lobärpneumonie Pasteurella multocida, Mycoplasma mycoides, Mannheimia hämolytica, Histophilus somni, Actinobacillus pneumoniae Bronchopneumonie Pasteurella multocida, Bordetella bronchiseptica, Trueperella pyogenes, Streptococcus spp., Escherichia coli Interstitielle Pneumonie Mykoplasmen, Influenza A/B, Adenoviren, Enteroviren, Respiratorisches Synzytialvirus A/B, Picornaviren, Paramyxoviren, Masernviren Embolisch-metastatische Pneumonie Trueperella pyogenes, Fusobacterium necrophorum, Erysipelothrix rhusiopathiae, Streptokokken und Staphylokokken Granulomatöse Pneumonie Mykobakterien (z. B. M. tuberculosis, M. bovis), Pilze (z. B. Blastomykose, Kryptokokkose), Parasiten (z. B. Filaroides spp., Pneumonyssus), Fremdkörper 2.2.6.3 Sonderformen Embolisch-metastatische Pneumonien Bei dieser Form der Pneumonie gelangen die Erreger nicht aerogen, sondern hämatogen in die Lunge. Während sterile Thromben durch Fibrinolyse aufgelöst werden, rufen septische Thromben eine Entzündungsreaktion der Lungenarterien und alveolären Kapillaren hervor, die sich in das angrenzende Interstitium ausbreitet. Makroskopisch ist die embolisch-metastatische Pneumonie durch multifokale, willkürlich im Lungengewebe verteilte, unterschiedlich große Abszesse charakterisiert. Beim Haustier sind als einige der häufigsten Ursachen Rupturen von Leberabszessen in die Vena cava caudalis, Omphalophlebitiden bei Neonaten oder Endokarditiden der rechten Herzseite beschrieben. Hier zählen Trueperella pyogenes, Fusobacterium necrophorum, Erysipelothrix rusiopathiae, Streptokokken und Staphylokokken zu den wichtigsten Erregern (MCGAVIN u. ZACHARY 2009). Beim Menschen sind ebenfalls Streptokokken und Staphylokokken beteiligt, aber 16 Literaturübersicht auch Meningokokken, Haemophilus influenzae, Pseudomonas aeruginosa und Proteus mirabilis (BÜHLING et al. 2004). 2.2.6.4 Granulomatöse Pneumonien Ursächlich für granulomatöse Pneumonien sind Organismen oder Fremdkörper, die resistent gegen intrazelluläre Lyse durch Phagozyten sind und im Lungengewebe eine lokale Entzündungsreaktion hervorrufen. Makroskopisch stellen sie sich als knotige, unterschiedlich große Gewebsveränderungen dar. Histologisch weisen die Granulome einen typischen Aufbau auf. So besteht zum Beispiel das in der Lunge anzutreffende Tuberkulosegranulom aus einem nekrotischen Zentrum, das von einem Saum aus Makrophagen (Epitheloidzellen) und Riesenzellen und von einer äußeren, von Lymphozyten und Plasmazellen infiltrierten Bindegewebsschicht umgeben ist. Abhängig vom auslösenden Agens und Zusammensetzung der Entzündungszellen kann der Granulomaufbau jedoch variieren. Zu den häufigsten Pathogenen bei Tieren zählen Mykobakterien, Pilze, Parasiten und Fremdkörper, welche entweder aerogen oder hämatogen in die Lunge gelangen. Beim Menschen sind vor allem Mycobacterium (M.) tuberculosis, seltener M. bovis und die Sarkoidose von Bedeutung. 2.2.7 Tumoren Im Gegensatz zum Menschen, bei dem das Rauchen den Hauptgrund für Lungentumoren darstellt, sind bei nicht humanen Primaten primäre Neoplasien in der Lunge selten (ABEE et al. 2012). Für Weißbüschelaffen liegt lediglich ein Fallbericht über ein bronchiales Adenom vor (BRACK et al. 1996). Weitere gutartige, epitheliale Neoplasien, wie das bronchio-alveoläre oder das bronchiale Adenom, sind beim Rhesus- und Javaneraffen (MILLER 1994) beschrieben. Ebenfalls bei Javaner(KASPAREIT et al. 2007) und Rhesusaffen (NEUSSER et al. 2001) liegen Berichte über maligne, epitheliale Neoplasien, wie z. B. ein bronchio-alveoläres Karzinom, ein Plattenepithelkarzinom und ein Karzinoid vor. Gutartige, mesenchymale Primärtumoren in der Lunge sind nicht publiziert, wohingegen das zu den malignen, mesenchymalen Tumoren gehörende pleurale Mesotheliom unter anderem beim 17 Literaturübersicht Anubispavian (Papio anubis) (FORTMAN et al. 1993) und Rhesusaffen (YAMATE et al. 2007) beschrieben wurde. Aufgrund der großen Anzahl von Kapillaren und Lymphgefäßen treten bei Haussäugetieren in der Lunge sekundäre, metastatische Tumoren deutlich häufiger auf als die primären Neoplasien (CASWELL u. WILLIAMS 2008). Auch bei nicht humanen Primaten ist bekannt, dass Lunge und Leber die Hauptzielorgane für hämatogene Metastasen darstellen (CABEE et al. 2012). Wenn auch nicht spezifisch für den Weißbüschelaffen, so ist bei verschiedensten Neu- und Altweltaffen eine Vielzahl von Tumormetastasen in der Lunge beschrieben. 2.3 Respiratorische Pathogene bei nicht humanen Primaten 2.3.1 Viren Virale Erreger des Respirationstrakts spielen vor allem bei katarrhalischen Entzündungen der Bronchien und Bronchiolen (akute Entzündung der Schleimhaut mit erhöhter Muzinsekretion) sowie bei interstitiellen Pneumonien eine Rolle. Bei nicht humanen Primaten und humanen Primaten zählen zu den wichtigsten Erregern das Respiratorische Synzytialvirus A und B (SZENTIKS et al. 2009), das Humane Metapneumovirus (SCHILDGEN et al. 2007), das Influenza A Virus (RIMMELZWAAN et al. 2001), das Influenza B Virus (KITANO et al. 2010), das Parainfluenzavirus (SUTHERLAND et al. 1986), Adenoviren (UMEMURA et al. 1985), Rhinoviren (DICK u. DICK 1974) und Masernviren (HALL et al. 1971). Das zur Familie der Paramyxoviridae gehörende Respiratorische Synzytialvirus (RSV) wurde ursprünglich 1955 aus dem Respirationstrakt eines Schimpansen (Pan sp.) isoliert, woraufhin kurze Zeit später die erste Infektion beim Menschen beschrieben worden ist. Seit seiner Entdeckung stellt es weltweit eine der wichtigsten infektiösen Ursachen für Lungenerkrankungen bei Säuglingen und Kindern dar (MCARTHUR-VAUGHAN u. GERSHWIN 2002). Das Virus ist hochkontagiös, wird über Aerosole verbreitet, und Anti-RSV Antikörper sind bei Menschen und nicht humanen Primaten weit verbreitet (RICHARDSON-WYATT et al. 1981). Unter den Altweltaffen sind Erkrankungen vor allem bei Schimpansen (KONDGEN et al. 2010), Indischen Hutaffen (Macaca radiata) (SIMOES et al. 1999) und juvenilen 18 Literaturübersicht Rhesusaffen (Macaca mulatta) (MCARTHUR-VAUGHAN u. GERSHWIN 2002) beschrieben, wohingegen Neuweltaffen wie Kapuziner- (Cebus) und Totenkopfaffen (Saimiri) sich zwar experimentell infizieren lassen, aber keine Symptome zeigen (BELSHE et al. 1977). Erstinfektionen bei Jungtieren betreffen den unteren Respirationstrakt und stellen sich oft als Bronchiolitis dar. Bei älteren Tieren, die schon vorher mit dem Erreger Kontakt hatten, ist die Infektion auf den oberen Respirationstrakt beschränkt (ABEE et al. 2012). Die Erkrankung verläuft selbstlimitierend, kann aber bakterielle Sekundärinfektionen begünstigen (GUSTAVSSON et al. 1990). Über Infektionen beim Weißbüschelaffen liegen keine Daten in der Literatur vor. Seit seiner Entdeckung 2001 stellt das ebenfalls zur Familie der Paramyxoviridae gehörende Humane Metapneumovirus (HMPV), eine sich weltweit ausbreitende, respiratorische Infektionserkrankung bei Kindern und immunsupprimierten Erwachsenen dar (SCHILDGEN et al. 2007; SHAHDA et al. 2011). Bei nicht humanen Primaten ist bisher noch wenig über Pathogenese und Verbreitung dieses Virus bekannt. Experimentell ließen sich Javaneraffen (KUIKEN et al. 2004), Rhesusaffen, Grüne Meerkatzen (Cercopithecus aethiops) (MACPHAIL et al. 2004) und Schimpansen (Pan troglodytes) (SKIADOPOULOS et al. 2004) infizieren, wobei die klinischen Symptome und pathomorphologischen Veränderungen einen deutlich geringeren Schweregrad aufwiesen als die des Menschen, bei dem die Erkrankung mit Bronchitiden und Pneumonien vergesellschaftet ist (SCHILDGEN et al. 2007). Als milde Rhinitiden mit Verlust des zilierten Epithels sowie minimale bis geringgradige, multifokale Erosionen in den Luft führenden Wegen und einer erhöhten Anzahl von Alveolarmakrophagen beschreibt KUIKEN (2004) die Infektion bei Javaneraffen. Nach SKIADOPOULOS et al. (2004) zeigten die experimentell infizierten Schimpansen klinische Symptome wie Husten. Dem Autor nach deutet die hohe Seroprävalenz für HMPV bei diesen Tieren darauf hin, dass eine natürliche Virusübertragung zwischen den Schimpansen sowie vom Menschen auf den Schimpansen möglich ist. Über Infektionen beim Weißbüschelaffen liegen keine Daten in der Literatur vor. 19 Literaturübersicht Bei Influenzavirus A und B handelt es sich um zu der Gruppe der Orthomyxoviridae gehörende, weltweit verbreitete, hochkontagiöse Viren, die durch Aerosole übertragen werden und respiratorische Erkrankungen bei Tier und Mensch hervorrufen. Übertragungen sind bei dieser Anthropozoonose nicht nur von Tier zu Tier, sondern auch vom Menschen auf den nicht humanen Primaten möglich. Infektionen treten gleichermaßen bei Altwelt- und Neuweltaffen auf, sind bei Weißbüschelaffen aber nicht dokumentiert. Milde Verläufen stellen sich als minimale Entzündungen des oberen Respirationstrakts dar, bestehend aus hyperämischen Schleimhäuten und erhöhter Mukussekretion. Schwerere Verläufe weisen eine Mitbeteiligung der intrapulmonalen Luftwege auf, wobei bakterielle Sekundärinfektionen auftreten können, die sich makroskopisch als katarrhalische oder mukopurulente Entzündung (vermehrte Muzin und Eiterbildung) von den Nasengängen bis in die Bronchiolen darstellen (MCGAVIN u. ZACHARY 2009; ABEE et al. 2012). Schwere Krankheitsverläufe sind bei Infektionen des Menschen und Javaneraffen mit Influenza A H5N1 und H1N1v beschrieben, die sich histopathologisch als hochgradige nekrotisierende, bronchointerstitielle Pneumonie (H5N1) oder diffuse Schädigung der Typ 1-Pneumozyten, des Endothels und der alveolären Septen (H1N1v) zeigen. Aufgrund der vergleichbaren pathomorphologischen Ausprägung dieser Erkrankung bei Mensch und nicht humanen Primaten dient hier der Javaneraffe als etabliertes Tiermodell (KUIKEN et al. 2003; HERFST et al. 2010). Anzeichen milder, entzündlicher, respiratorischer Veränderungen, aber auch schwerwiegende Rhinotracheobronchitiden und interstitielle Pneumonien finden sich bei Parainfluenzavirus-Infektionen des Weißbüschelaffen. Von diesen zur Gattung der Paramyxoviren gehörenden Erregern sind vor allem Typ 1, 2 und 3 mit Erkrankungen bei Neuweltaffen assoziiert, wobei bei Weißbüschelaffen vor allem Infektionen mit dem Typ 1 auftreten. Histopathologisch ist diese Infektion durch den Nachweis von mehrkernigen intrazytoplasmatischen Synzytialzellen Einschlusskörperchen mit intranukleären gekennzeichnet. Auch und beim Menschen, vor allem bei Säuglingen und Kindern, sind Parainfluenzavirus Typ 1, 2 und 3 eine häufige Ursache oberer und unterer Respirationstrakterkrankungen 20 Literaturübersicht (FLECKNELL et al. 1983; SUTHERLAND et al. 1986; POTKAY 1992; DURBIN et al. 2000; LUDLAGE u. MANSFIELD 2003). Die Familie der Adenoviridae setzt sich aus einer großen Anzahl ubiquitärer, weltweit verbreiteter Viren zusammen, die als sporadische sowie endemische Infektion bei Menschen und Wirbeltieren auftritt. Insbesondere bei Säuglingen, Kleinkindern, Immunsupprimierten und Transplantatempfängern sind letale, disseminierte Krankheitsverläufe dieser Infektion beschrieben, die sonst vor allem als selbst-limitierende Erkrankung des Respirationstrakts verläuft (LOUIE et al. 2008; CALCEDO et al. 2009). Auch bei nicht humanen Primaten sind klinische Manifestationen, vor allem im Respirations- und Gastrointestinaltrakt, bei neugeborenen, juvenilen oder immunsupprimierten Tieren dokumentiert, inapparente Verläufe dagegen bei adulten, immunkompetenten Tieren. Neben der oro-nasalen Infektionsroute durch Aerosole stellt die fäkal-orale Transmission ebenfalls einen bedeutenden Infektionsweg dar. Dieser wird durch eine persistierende Infektion in lymphatischem Gewebe und intestinalem Epithel ermöglicht und geht mit einer Erregerausscheidung einher, selbst lange nach Abklingen der klinischen Symptome. Makroskopisch zeigt sich die Lunge oftmals deutlich vergrößert mit multifokalen, verdichteten Bereichen. Histopathologisch ist dieses Krankheitsbild durch eine nekrotisierende Alveolitis und Bronchiolitis gekennzeichnet mit großen, basophilen, intranukleären Einschlusskörperchen (BOYCE et al. 1978; ABEE et al. 2012). Adenoviren wurden bei Krallenaffen, Weißbüschelaffen, Roten Springaffen (Callicebus cupreus) und Rotbauchtamarinen (Saguinus labiatus) nachgewiesen, aber auch bei diversen, in Gefangenschaft und freier Wildbahn lebenden Altwelt- und Menschenaffen (BOYCE et al. 1978; CHEN et al. 2011; WEVERS et al. 2011). Beim Menschen wurden Infektionen mit dem Humanen Rhinovirus (HRV) bereits vor 50 Jahren festgestellt. Aber erst durch klinische Studien und mithilfe der Entwicklung neuer molekularbiologischer Methoden konnte belegt werden, dass HRV-Infektionen die zweithäufigste Ursache für Pneumonien und Bronchiolitiden bei Neugeborenen und Kleinkindern darstellen, und dass sie an progressiven Verläufen von präexistenten respiratorischen Erkrankungen wie Asthma, chronisch obstruktive Lungenerkrankung (COPD) und zystischer Fibrose beteiligt sind (HAYDEN 2004; 21 Literaturübersicht GERN 2010). Aufgrund eines ausgeprägten Zelltropismus des Virus sind nach XATZIPSALTI und PAPADOPOULOS (2007) neben dem Schimpansen nur noch Gibbons (Hylobates sp.) für humane Rhinoviren empfänglich. Experimentelle und natürliche Infektionen sind bei Schimpansen beschrieben, wobei die Erkrankung oft klinisch inapparent oder mild verläuft (DICK u. DICK 1974). Gibbons stellen sich als weniger empfänglich dar. Klinische Symptome sowie Läsionen sind nicht beschrieben. Inokulationen bei Neuweltaffen und Totenkopfaffen (Saimiri sciureus) sowie Weißstirnkapuziner (Cebus albifrons) blieben erfolglos (ABEE et al. 2012). Masernviren, die unter anderem mit dem kaninen Staupevirus und dem Rinderpestvirus die Gattung der Morbilliviren (Paramyxoviridae) bilden, infizieren eine große Anzahl von nicht humanen Primaten. Während das Virus bei Altweltaffen ausgeprägte kutane Veränderungen sowie Entzündungen des Respirationstraktes hervorruft, verursacht es bei Krallenaffen nekrotisierende Enteritiden mit hämorrhagischer Diarrhoe. Weniger häufig treten nekrotisierende, interstitielle Pneumonien auf. Schwere Krankheitsverläufe mit hohen Morbiditäts- und Mortalitätsraten sind beschrieben, die vor allem bei immunsupprimierten Tieren bis zu 100 % erreichen können. Eine Verdachtsdiagnose kann auf der Grundlage der klinischen Symptome in Verbindung mit dem histologischen Nachweis einer Enteritis mit mehrkernigen Riesenzellen und basophilen Einschlusskörperchen gestellt werden (ALBRECHT et al. 1980; LUDLAGE u. MANSFIELD 2003; BAILEY u. MANSFIELD 2010; ABEE et al. 2012). 2.3.2 Bakterien Bakterielle Pathogene sind im Respirationstrakt an verschieden Ausprägungsformen der Pneumonie beteiligt. Bei nicht humanen Primaten sind zahlreiche bakterielle Infektionserreger beschrieben, die zu einem breiten Spektrum an Lungenveränderungen führen können. Abhängig vom Agens und dem Ausmaß des Gewebsschadens entwickeln sich alveoläre Herdpneumonien, wie die katarrhalischeitrige Bronchopneumonie oder die fibrinöse Pneumonie. Ebenfalls können Sonderformen wie die granulomatöse oder die embolisch-metastatische Pneumonie 22 Literaturübersicht auftreten. Im Folgenden sind die häufigsten bakteriellen Erreger des unteren Respirationstrakts bei nicht humanen Primaten aufgelistet. Bordetella bronchiseptica und Klebsiella pneumoniae gehören zu den relevanten Pathogenen des Respirationstraktes beim Weißbüschelaffen. Die Infektion mit Bordetellen erfolgt in der Regel über Aerosole. Drastische Krankheitsverläufe und hohe Mortalitätsraten werden vor allem bei Tieren unter einem Jahr beschrieben. Klebsiella pneumoniae, ein Saprophyt des Nasopharynx und Gastrointestinaltrakts des Menschen, ist beim Weißbüschelaffen als Erreger von Pneumonien und Enteritiden bekannt und spielt vor allem bei immunsupprimierten Tieren eine Rolle (LUDLAGE u. MANSFIELD 2003). Bei Streptococcus equi ssp. zooepdemicus handelt es sich um einen opportunistischen Erreger, der bei Pferden zur normalen Flora der oberen Atemwege zählt und bei geschwächter Abwehrlage, z. B. als Folge einer primären Virusinfektion, sekundäre bakterielle Bronchopneumonien hervorrufen kann (MCGAVIN u. ZACHARY 2009). Nicht humane Primaten, vor allem Rhesusaffen, scheinen hoch empfänglich für diesen Erreger zu sein. Infektionen verlaufen hier oft tödlich und bleiben nicht auf immunsupprimierte Tiere beschränkt (MÄTZ-RENSING et al. 2009). Neben septikämischen Verläufen bei Rotbauch- und Springtamarinen (SCHILLER et al. 1989) sind Enteritiden bei Makaken, Meerkatzen und dem Mandrill (Mandrillus sphinx) (DZHIKIDZE et al. 1996) bekannt. Die von MÄTZ-RENSING et al. (2009) beschriebenen Infektionen beim Rhesusaffen zeigten sich histopathologisch als purulente Konjunktivitis, Rhinitis, Pharyngitis und hochgradige fibrinopurulente Bronchopneumonie. Die Entzündung breitete sich ebenfalls auf die angrenzenden Organe wie Herz und Diaphragma aus. Übertragen wird Streptococcus equi über Aerosole direkt von Tier zu Tier, wobei auch Transmissionen zwischen Tier und Mensch möglich sind (BRACK et al. 1997; MÄTZ-RENSING et al. 2009). Orale Infektionen sind aus einer Krallenaffenkolonie bekannt, die sich vermutlich über infiziertes Pferdefleisch angesteckt hatte, das an im selben Gehege gehaltene Gürteltiere verfüttert wurde (SCHILLER et al. 1989). Streptococcus pneumoniae zählt zu den Kommensalen des oberen Respirationstraktes beim Menschen. Bei immunsupprimierten Personen ist er an 23 Literaturübersicht einer Vielzahl von Erkrankungen wie Meningitis, Otitis sowie Sinusitis beteiligt und stellt außerhalb des Krankenhauses (ambulant erworbene Pneumonie) den häufigsten Pneumonieerreger dar (PHILIPP et al. 2006; ZOU et al. 2010). Im Rhesusaffen, Schimpansen und Totenkopfaffen sind Streptokokken mit multifokalen Bronchopneumonien, Abszessen und Atelektasen vergesellschaftet, wobei die Tiere entweder experimentell infiziert wurden oder in direktem Kontakt zum Menschen standen, was die Vermutung nahe legt, dass der Mensch als Vektor fungiert (BERENDT et al. 1975; PHILIPP et al. 2006; CHI et al. 2007; ZOU et al. 2010). Pasteurella multocida und Pasteurella haemolytica sind beim Tier an dem Krankheitsbild der Pasteurellose beteiligt, die als perakute oder akute Septikämie verläuft, wobei deren Symptome von dem betroffenen Organsystem abhängen. Immunsupprimierende Primärinfektionen und Stress, z. B. hervorgerufen durch Transport oder Überbelegung, wirken sich prädisponierend auf eine Infektion aus. Eine Manifestation dieser Erkrankung im Respirationstrakt, die sich histopathologisch als fibrinopurulente oder interstitielle Pneumonie mit Fibrinthromben und Bakterien in pulmonalen Gefäßen zeigt, ist beim Springtamarin (DUNCAN et al. 1995), Östlichen Graukehl-Nachtaffen (Aotus trivirgatus) (BENJAMIN u. LANG 1971) und Husarenaffen (Erythrocebus patas) (OKOH u. OCHOLI 1986) bekannt. Die Lungentuberkulose zählt zu den granulomatösen Pneumonien und wird im Primaten wie im Menschen durch Mycobacterium bovis und Mycobacterium tuberculosis hervorgerufen. Da sich das Krankheitsbild bei beiden Spezies kaum voneinander unterscheidet, dient der Javaneraffe (Macaca fascicularis) als etabliertes Tiermodell in der Tuberkuloseforschung (FLYNN et al. 2003). Die Infektion erfolgt in den meisten Fällen über die aerogene Aufnahme infizierter Aerosole, seltener auf oralem Weg, wobei der Übertragung vom Menschen auf den Primaten eine besondere Bedeutung zukommt und Infektionen in abgeschiedenen Wildpopulationen kaum auftreten. Makroskopisch zeichnet sich die Tuberkulose durch verkäsende, teils konfluierende Granulome aus, die sich lymphogen im Lungengewebe ausbreiten und im fortgeschrittenen Stadium in Leber, Niere, Milz und verschiedenen Lymphknoten anzutreffen sind. Histologisch variiert die Ausprägung von typischen Granulomen mit nekrotischem Zentrum, umgeben von 24 Literaturübersicht epitheloiden Makrophagen, Langhans-Riesenzellen und Lymphozyten bis zu wenig organisierten Pyogranulomen. Säurefeste Bakterien lassen sich gelegentlich im Zentrum des Granuloms oder in dessen Peripherie nachweisen. Im chronischen Stadium nimmt der Anteil von Makrophagen und Riesenzellen ab, und die Granulome können dystrophisch verkalken. Unter den Altweltaffen scheinen besonders Rhesus- und Javaneraffen für Tuberkulose empfänglich zu sein. Bei Neuweltaffen sind nur sporadische Infektionen bei Totenkopfaffen (HESSLER u. MORELAND 1968), Kapuzineraffen (LEATHERS u. HAMM 1976) und Östlichen Graukehl-Nachtaffen beschrieben (SNYDER et al. 1970; HUNT et al. 1978; BAILEY u. MANSFIELD 2010). 2.3.4 Parasiten Protozoen Infektionen mit Toxoplasma gondii treten bei Halb-, Altwelt- und Neuweltaffen auf (JONES et al. 1993) und verlaufen bei Krallen- und Halbaffen meist tödlich. Beim Altweltaffen hingegen handelt es sich oftmals um selbst-limitierende, klinisch inapparente Erkrankungen, die im Zusammenhang mit Immunsuppressionen festgestellt werden. Die Übertragung von Toxoplasmen erfolgt entweder über die direkte oder indirekte Aufnahme sporulierter Oozysten, die von der Katze als Endwirt mit dem Kot ausgeschieden werden oder über Nagetiere, die als Zwischenwirte infektiöse Gewebszysten enthalten (HILL u. DUBEY 2002). Klinische Symptome sind oft unspezifisch wie z. B. Anorexie, Lethargie, Husten und Nasen- sowie Augenausfluss. Mikroskopisch lassen sich in verschiedenen Organen multifokale Nekrosen durch sich schnell teilende Tachyzoiten nachweisen. Bei einer Manifestation im Respirationstrakts zeigen sich histopathologisch Nekrosen des Gefäßendothels sowie als Folge der regenerativen Prozesse im Alveolarbereich eine Typ 2-Pneumozyten-Hyperplasie (ABEE et al. 2012). Zestoden Nicht humane Primaten können als Zwischenwirt für eine Vielzahl von Zestoden dienen, wobei aufgrund ihres mitunter destruktiven Charakters insbesondere Taenia 25 Literaturübersicht spp. und Echinococcus spp. in der Lunge als auch in anderen Organen erheblichen Schaden anrichten können. Erkrankungen sind bei Altwelt-, Halb- und bei Neuweltaffen beschrieben. Fallberichte für Lungenmanifestationen bei Weißbüschelaffen liegen nicht vor (KONDO et al. 1996; TAPPE et al. 2007; TSUBOTA et al. 2009; LUZON et al. 2010; BORJI et al. 2012). Nematoden Pulmonale Nematodiose, hervorgerufen durch Filaroides spp. und Filariopsis spp., wurden bei verschieden Alt- und Neuweltaffen beschrieben. Neben Marmosetten sind Infektionen ebenfalls bei Totenkopfaffen, Kapuzineraffen und Brüllaffen (Alouatta sp.) beschrieben (GARCIA et al. 2009). Die adulten Metastrongyliden sitzen in den terminalen Bronchiolen und Alveolen, produzieren Larven, welche hochgehustet, abgeschluckt und mit den Fäzes ausgeschieden werden. Makroskopisch stellen sich in der Lunge kleine, subpleurale, pink-graue Herde dar, welche histologisch interstitielle Fibrose, gemischtzelliges Entzündungszellinfiltrat und Epithelveränderungen wie Atrophie, Hyperplasie als auch Metaplasie aufweisen können (ABEE et al. 2012). Weitere bedeutende Parasiten bei Neu- und Altweltaffen sind Gongylonema macrogubernaculum und Gongylonema puchrum. Diese weltweiltverbreiteten Errerger, finden sich bei infizierten Tiere vor allem im Ösophagus, Lippen, Maulschleimhaut, Zunge, Magen und Bronchien. Während sie in der Regel mit asymptomatischen Krankheitsverläufen beschrieben worden sind, konnte kürzlich ein vermehrtes Auftreten von Ösophaguskarzinomen mit diesem Erreger in Verbindung gebracht werden (BLEIER et al. 2005; ABEE et al. 2012) Arthropoden Die Erreger der pulmonalen Acariasis sind bei Altweltaffen Lungenmilben der Gattung Pneumonyssus. Eine ähnliche Spezies, Pneumonyssoides, verursacht bei Neuweltaffen vergleichbare Läsionen, welche makroskopisch mit wenigen Millimeter großen, runden, weiß-gelben Herden einhergehen, in denen eine oder mehrere Milben sitzen. Histologisch zeichnen sich diese Veränderungen durch dilatierte und verdickte Bronchiolen aus, in denen sich das respiratorische Epithel teilweise ablöst 26 Literaturübersicht und gemischtzellige Entzündungsinfiltrationen auftreten. Die Makrophagen enthalten oftmals phagozytiertes, kristallines, goldbraunes Parasitenpigment (KIM u. KALTER 1975; MCCLURE et al. 1976; ANDRADE u. MARCHEVSKY 2007; ABEE et al. 2012). 2.3.3 Pilze Zu den Systemmykosen, die in der Regel granulomatöse Pneumonien im nicht humanen Primaten auslösen, zählt die Kokzidiomykose, die Histoplasmose, die Kryptokokkose und die Blastomykose. Der Erreger der Kokzidiomykose Coccidioides immitis ist auf dem amerikanischen Kontinent endemisch (LEE et al. 2008). Aus dem Myzel dieses dimorphen Schlauchpilzes entwickeln sich nach Regenfällen Arthrokonidien, die aerogen aufgenommen werden und sich in der Lunge zu parasitär wachsenden Hefen entwickeln und granulomatöse bis pyogranulomatöse Pneumonien verursachen (CASWELL u. WILLIAMS 2008). Spontane Infektionen sind unter anderem bei Schimpansen (HERRIN et al. 2005) und Japanmakaken (Macaca fuscata) beschrieben (GRAYBILL et al. 1990). Für den Weißbüschelaffen liegen keine Fallbeschreibungen vor. Bei der Histoplasmose, hervorgerufen durch Histoplasma capsulatum, handelt es sich ebenfalls um einen dimorphen Pilz. In der Umwelt wächst Histoplasma capsulatum als nicht parasitäre Myzelform. Infektionen erfolgen durch aerogene Aufnahme der Sporen, die im Lungengewebe auskeimen und parasitäre Hefeformen bilden. Erkrankungen mit diesem Pilz sind bei einer Vielzahl von Tieren bekannt (KUROWSKI u. OSTAPCHUK 2002; CASWELL u. WILLIAMS 2008). Neben Fallbeschreibungen beim Altweltaffen (COSTA et al. 1994; WACHTMAN u. MANSFIELD 2008), liegen ebenfalls Berichte für Spix-Nachtaffen (Aotus vociferans) (MILLER u. OWENS 1999) und WeißkopfBüschelaffen (Callithrix geoffroyi) (DA CRUZ et al. 1993) vor. Beim Weißbüschelaffen liegen keine Erkenntnisse über Erkrankungen an Histoplasmose vor, jedoch gelang COSTA et al. (1994) mithilfe von intradermalen Hypersensitivitätstests der Nachweis von Infektionen mit Histoplasma capsulatum bei dieser Tierart. Die Kryptokokkose tritt weltweit als eine opportunistische Infektion auf, die aerogen über den im Erdreich lebenden Pilz Cryptococcus neoformans 27 Literaturübersicht übertragen wird und zu granulomatösen Entzündungen in Nase und Lunge führt mit disseminierter Erregerausbreitung, wobei der Pilz im Menschen eine Prädisposition für das zentrale Nervensystem aufweist. Erkrankungen sind beim Altwelt- (PAL et al. 1984) sowie beim Neuweltaffen (ROUSSILHON et al. 1987) beschrieben. JUANSALLÉS et al. (1998) dokumentierten eine intestinale Verlaufsform der Kryptokokkose beim Weißbüschelaffen, was vermuten lässt, dass neben der bekannten aerogenen Infektionsroute ebenfalls eine orale Übertragung möglich ist. Bei dem dimorphen Pilz Blastomyces dermatitidis, dem Erreger der Blastomykose, produziert dessen Myzelform Sporen, die vom Wirt aerogen aufgenommen werden. In der Lunge lassen sich multifokale, grau-weiße Knoten feststellen. Histologisch weisen diese Veränderungen einen typischen Granulomaufbau auf, bestehend aus epitheloiden Makrophagen, Riesenzellen, neutrophilen Granulozyten, Lymphozyten und Bindegewebe. Die Hefen sind im Durchmesser 5-15 µm groß, rund und nicht bekapselt, besitzen jedoch eine ca. 1 µm dicke Zellwand (CASWELL u. WILLIAMS 2008). Empfänglich ist neben dem Menschen eine Vielzahl von Tieren. Bei nicht humanen Primaten liegen Fallbeschreibungen einer Blastomykose beim Makaken (WILKINSON et al. 1999) und Nilgiri-Languren (Trachypithecus johnii) vor (CHAKRABORTY u. GOSWAMI 1996). Bei Weißbüschelaffen sind keine Fälle von Blastomykose bekannt. Mithilfe molekularbiologischer Methoden wurde von EDMAN (1988) festgestellt, dass es sich bei denen in der Vergangenheit als Protozoon eingestuften Pneumocystis spp. um Pilze handelt. Bei immunsupprimierten Menschen als Pneumocystis jirovecii bekannt, sind Altwelt- und Neuweltaffen empfänglich für Pneumocystis carinii. Im Gegensatz zu anderen Pilzen verläuft die Pneumozystose nicht als granulomatöse, sondern als chronisch interstitielle Pneumonie. Dieser opportunistische Pilz tritt in der Lunge entweder als Trophozoit oder Zyste auf. Die dünnwandigen Trophozoiten sind 1-4 µm groß und besitzen, im Gegensatz zu den mehrkernigen, dickwandigen, 5-8 µm großen Zysten, einen Zellkern. Makroskopisch fallen rot-braune, multifokalkonfluierende, verhärtete Bereiche auf. Neben der interstitiellen Pneumonie tritt als typische histologische Veränderung schaumiges Material in den Alveolen auf, bestehend aus rund- bis sichelförmigen Pilzstrukturen (CASWELL u. WILLIAMS 28 Literaturübersicht 2008; WACHTMAN u. MANSFIELD 2008; ABEE et al. 2012). Infektionen sind vor allem bei Menschen, die mit dem humanen Immundefizienz-Virus (ZHIVOTOVSKAIA) infiziert sind, bei simianen Immundefizienz-Virus (SIV) infizierten Makaken (YANAI et al. 1999) und bei Araberfohlen mit „severe combined immunodeficiency“ (SCID) (PERRYMAN 2004), beschrieben. Bei Krallenaffen liegen Fallberichte beim Braunrückentamarin (Saguinus fuscicollis) und Lisztaffen (Saguinus oedipus) (RICHTER et al. 1978) vor. Für den Weißbüschelaffen sind keine Infektionen mit Pneumocystis carinii publiziert. 29 Eigene Untersuchungen 3 Eigene Untersuchungen 3.1 Material und Methoden 3.1.1 Allgemeine Daten Die Grundlage dieser Arbeit bilden teils retrospektive, teils aktuell erfasste Daten von insgesamt 700 Weißbüschelaffen aus dem Sektionsgut der Abteilung Infektionspathologie des Deutschen Primatenzentrums (DPZ) in Göttingen, die in dem Zeitraum von 1997 bis 2011 gesammelt wurden. Der überwiegende Anteil der Tiere stammt aus der eigenen Kolonie des Deutschen Primatenzentrums und bildet den Schwerpunkt dieser Arbeit. Bei einem kleineren Teil handelt es sich um Weißbüschelaffen aus externen Forschungs- bzw. Laboreinrichtungen. Durch abweichende Haltungs- und Fütterungsbedingungen in den verschiedenen Instituten erfolgte eine Gruppierung der Tiere in Untersuchungsgruppe I (n = 639), welche die Tiere des DPZ umfasst, und Untersuchungsgruppen II (n = 38) und III (n = 34), bestehend aus Tieren aus zwei externen Kolonien. Diese setzen sich zum einen aus Tieren der Universität Göttingen und zum anderen aus Tieren eines Pharmaunternehmens zusammen. Die Auswertung der Protokolle, anhand derer jede Sektion dokumentiert wird, ermöglichte die Ermittlung der Sektionsgesamtzahl und die Erhebung tierartspezifischer Informationen wie Alter und Geschlecht. 3.1.2 Sektion und Probenentnahme Bei denen in die vorliegende Studie aufgenommen Weißbüschelaffen handelte es sich um Tiere, die entweder verstorben sind, oder die krankheitsbedingt bzw. im Rahmen experimenteller Arbeiten euthanasiert werden mussten. Hierfür wurden sie zunächst mit der „Göttinger Mischung II“ (Anhang 9.1) narkotisiert und anschließend mit einer überdosierten Injektion Pentobarbital-Natrium (Narcoren®; ca. 150 mg/kg KGW) intraperitoneal oder intrakardial euthanasiert. Der Sektionsgang erfolgte nach einem standardisierten Ablauf, welcher die detaillierte Beschreibung aller makroskopischen Befunde und die Entnahme eines umfangreichen Organspektrums für histologische Zwecke umfasste. Lungengewebe wurde zusammen mit den Proben aus verschiedenen Organsystemen in 10%igem neutral gepuffertem Formalin fixiert (Anhang 9.2). Im bakteriologische und in Rahmen der Sektion wurden Verdachtsfällen 30 parasitologische kulturell- Untersuchungen Eigene Untersuchungen durchgeführt. Zusätzlich wurde seit dem Jahr 2011 bei aktuellen Fällen Lungengewebe entnommen, in Flüssigstickstoff eingefroren und bei -80 °C kryokonserviert, um diese mithilfe molekularbiologischer Methoden auf relevante Krankheitserreger zu testen. 3.1.3 Histologische Aufarbeitung Die entnommenen Gewebeproben wurden für mindestens 24 Stunden in 10%igem neutral gepuffertem Formalin bei Raumtemperatur fixiert. Nach dem Zuschneiden auf die Größe der Einbettkassetten erfolgte die automatische Einbettung (Shandon Excelsior ES, Fa. Thermo Scientific, Dreieich) in Paraffin nach laborüblichem Protokoll (Anhang 9.3). Anschließend wurden die Proben unter Verwendung einer Paraffinausgießstation (EC 350-1, Fa. Thermo Fisher Scientific, Dreieich) in passenden Stahlblechformen ausgegossen und mithilfe des Kassettenbodens aufgeblockt. Die ausgehärteten Blöcke wurden aus der Form gelöst und im Eisbad abgekühlt. Mithilfe eines Schlittenmikrotoms (Mikrotom HM 400R, Fa. Microm, Walldorf) erfolgte anschließend die Anfertigung von ca. 3 μm dicker Schnitten. Diese wurden mit einem zuvor in Eiswasser gekühlten Streifen Durchschlagpapier abgenommen, zur faltenfreien Ausdehnung in ein 40 °C warmes Wasserbad überführt und auf Standardobjektträger übertragen. Für die retrospektiven Untersuchungen wurde auf archiviertes Formalin-, Paraffin- und Schnittmaterial zurückgegriffen, das auf gleiche Weise aufgearbeitet worden war. Die angefertigten Paraffinschnitte des zu untersuchenden Organspektrums wurden im Färbeautomaten (Varistain Gemini, Fa. Thermo Shandon, Frankfurt am Main) mit Hämatoxilin-Eosin gefärbt (Anhang 9.4). Bei Bedarf wurden Berliner-Blau-Reaktionen zum Nachweis von dreiwertigen Eisenionen (Anhang 9.5) und die von Kossa-Färbung zum Nachweis von Kalzium und Kalziumsalzen (Anhang 9.6) durchgeführt. Des Weiteren erfolgten in Einzelfällen eine Trichromfärbung nach Masson-Goldner (Anhang 9.7) zur besseren Darstellung der mesenchymalen Gewebestrukturen sowie eine Kongorot-Färbung (Anhang 9.8) zum Nachweis möglicher Amyloidablagerungen. Pilzhyphen ließen sich mit der Versilberung nach Grocott (Anhang 9.9) und Bakterien 31 Eigene Untersuchungen sowohl mit einer Periodic-Acid-Schiff-Reaktion als auch mit einer Giemsa-Färbung darstellen (Anhang 9.10). Immunhistochemische Untersuchung der Entzündungszellen Um bei einer ausgewählten Tiergruppe mit interstitiellen Veränderungen die mononukleären Infiltrate in den Alveolarsepten auf ihren Anteil an B- und TLymphozyten zu untersuchen, wurde in Paraffin eingebettetes Lungengewebe immunhistochemisch mit anti-CD79- und anti-CD3-Antikörpern behandelt. Weiterhin wurden Makrophagen mit einem anti-MAC 387-Antikörper markiert. Die Tiergruppe, die in diese Untersuchung einbezogen wurde, setzte sich aus zwei juvenilen Weibchen und zwei juvenilen Männchen zusammen, wobei alle Tiere die am häufigsten vertretende Pneumonieform der gering- bis mittelgradigen, subakuten, multifokal-konfluierenden interstitiellen Entzündung aufwiesen. Die Auswertung erfolgte quantitativ im Lichtmikroskop mittels Handzählgerät, wobei zehn Gesichtsfelder lichtmikroskopisch bei vierzigfacher Vergrößerung mäanderförmig abgefahren wurden. Immunhistochemische Untersuchung von Bindegewebe und glatter Muskulatur Um gelegentlich auftretende pulmonale Gewebsproliferationen bei Tieren mit und ohne besonderen Befund in der Lunge weiter zu differenzieren, wurden diese immunhistochemisch mit anti-Vimentin- und anti-smooth muscle actin-Antikörpern markiert (Discovery XT Färbemodul, Fa. Roche, Basel; Anhang 9.11). 3.1.4 Mikrobiologische und parasitologische Untersuchungen Um die mögliche Beteiligung von Erregern am Krankheitsgeschehen nachzuweisen, wurden im Verdachtsfall eine parasitologische und routinemäßig eine kulturellbakteriologische Untersuchung durchgeführt. Der Nachweis von Parasiten erfolgte aus Dünn- und Dickdarmkot, welcher mit Wasser vermischt wurde, um diesen auf einem Objektträger auszustreichen. Ließen sich lichtmikroskopisch protozoäre Erregerstrukturen darstellen, wurde zur besseren morphologischen Beurteilung und Identifizierung eine Färbung mit Methylenblau oder Lugolscher Lösung durchgeführt 32 Eigene Untersuchungen (Anhang 9.12). Für die kulturell-bakteriologische Untersuchung wurden unmittelbar nach Eröffnung des Tierkörpers mit abgeflammtem Sektionsbesteck Inzisionen in Milz, Leber, Niere, Dünndarm, Dickdarm, Lunge und Herz und Großhirn gesetzt. Nach jeder Inzision erfolgte ein Abstrich des Organparenchyms unter Verwendung einer sterilen Einmal-Impföse, die daraufhin auf Blutagarplatten ausgestrichen wurde. Rachen- und Nasenabstriche wurden ebenfalls auf Blutagarplatten ausgestrichen. Um das intestinale komplette Keimspektrum zu erfassen, wurden des Weiteren Campylobacter- und Mac Conkey-Agar-Platten beimpft und ein SalmonellenAnreicherungsmedium angesetzt (Anhang 9.13). 3.1.5 Molekularbiologische Untersuchungen Mithilfe molekularbiologischer Methoden sind insgesamt 22 ausgewählte Proben von Tieren mit und ohne entzündliche Lungenveränderungen auf virale Krankheitserreger des Respirationstraktes untersucht worden. Zu diesen zählten Respiratorisches Synzytialvirus A und B, Influenza A, Influenza B, Adenovirus, Enterovirus, Picornaviren, Pan Paramyxoviren und Masern. Das Ausgangsmaterial stellte bei -80 °C asserviertes Lungengewebe dar, welches während der Sektion entnommen wurde. 3.1.5.1 RNA-Isolierung und Bestimmung der optischen Dichte Die Isolierung der Ribonukleinsäure (RNA) erfolgte mittels NucleoSpin RNA II (Fa. Macherey-Nagel, Düren, Anhang 9.14). Zu Beginn wurde ca. 60 µg Lungengewebe unter Zugabe von RA1 Puffer und β-Mercaptoethanol im 1,5-ml-Eppendorfcup mit einem sterilen Pistill mechanisch homogenisiert und das lysierte Gewebe daraufhin mittels NucleoSpin® Filter gereinigt. Die anschließende Zugabe von Ethanol ermöglichte die Bindung von RNA an die Silica-Membran. Nachdem sich im nächsten Schritt die RNA an die Silica-Membran angelagert hatte, wurde diese mittels MDB-Puffer entsalzt und noch vorhandene Desoxyribonukleinsäure (DNA) mit DNAse abgebaut. Die enzymatische DNA-Spaltung wurde durch den RA2 Puffer gestoppt und die Silica-Membran drei Mal gewaschen. Die nun vorhandene RNA wurde im letzten Schritt unter Zugabe von RNase freiem Wasser herausgelöst. 33 Eigene Untersuchungen Die Konzentrationsbestimmung der RNA-Lösung erfolgte photometrisch bei 260 nm (NanoDrop, Fa. Peqlab, Fürth). Für die Qualitätsbewertung wurden die Bereiche 230 und 280 nm miteinbezogen. 3.1.5.2 Reverse Transkription Um die RNA in die Polymerase-Kettenreaktion (PCR) einzusetzen, musste sie zuerst mittels einer Reversen Transkriptase (Kit Omniscript, Fa. Qiagen, Hilden) in komplementäre Desoxyribonukleinsäure (cDNA) umgeschrieben werden. Es wurden jeweils 500 ng pro Umschreibung eingesetzt. Um etwaige DNA-Kontaminationen zu detektieren, ist zu jeder Umschreibung auch eine RNA-Probe ohne Zugaben des Enzyms mitgeführt worden. Für die cDNA-Synthese wurde ein Oligo p(DT)15 Primer, der am Poly-A-Schwanz bindet, verwendet (Fa. Hoffmann-La Roche, Basel) (Anhang 9.15). 3.1.5.3 Quantitative Echtzeit-PCR (qRT-PCR) In der Real-Time PCR diente der Fluoreszenzfarbstoff SYBR Green als Detektionssystem. Die Spezifitätsprüfung erfolgte über die Schmelzkurvenanalyse. Primer: Zum spezifischen Nachweis der Ziel-cDNA mussten die eingesetzten Primer komplementär zur cDNA-Vorlage sein. Dies setzte voraus, dass die cDNA-Sequenz des entsprechenden Gens bekannt war. Die in dieser Dissertation verwendeten Primer waren zwischen 21 und 26 Nucleotide lang. Aufgrund der spezifischen Struktur lag die Annealing-Temperatur bei 60°C. Bei den verwendeten Primern handelte es sich um folgende: Respiratorisches Synzytialvirus (RSV): RSV-N15-foward (5’-GATGGCTGTTAGCAAAGTCAAGTT-3’) RSV-184-reverse (5’-CATCTTCWGTGATTAATARCATRCCACATA-3’) (REICHE u. SCHWEIGER 2009) 34 Eigene Untersuchungen Influenza A: M+25 (5’-AgATgAgTCTTCTAACCgAggTCg-3’) M-124BB (5’-ccWgCAAARACATCYTCAAgTYTCTg-3’) M-124sw (5’-CTgCAAAgACACTTTCCAgTCTCTg-3’) (SCHULZE et al. 2010) Influenza B: BMP-13 (5’-gAgACACAATgCCTACCTgC-3’) BMP-102AN (5’-TTCCCACCRAACCARCARTgTAAT-3’) (SCHULZE et al. 2010) Adenovirus: AD-024S (5’-GACGCYTCGGAGTACCTGAG-3’) AD-024R (5’-RGCCAGIGTRWAICGMRCYTTGTA-3’) (CHMIELEWICZ et al. 2005) Enterovirus: E-TM 1 S (5’-GCCCCTGAATGCGGCTAAT-3‘) E-TM 2 R (5‘-RATTGTCACCATAAGCAGYCA-3‘) (PUSCH et al. 2005) PCR-Bedingungen: Die Vorbereitung des PCR-Ansatzes erfolgte unter der Sterilbank. Der verwendete Mastermix ist in Tab. 2 aufgelistet. Er hatte ein Volumen von 25 µl. 35 Eigene Untersuchungen Tab. 2: Pipettierschema für die SYBR Green Real-Time PCR. Substanz Konzentration Menge H2O Endkonzentration 5 µl 2x ImmoMix 2x 12,5 µl 1x Magnesium 6 nmol SYBR Green 50x 1 µl 2x Rox 50x 0,5 µl 1x Primer, forward 10 pmol 0,5 µl 10 pmol Primer, reverse 10 pmol 0,5 µl 10 pmol 3 nmol Summe 20 µl Template 1:5 5 µl Gesamtvolumen 25 µl Bei jedem Lauf wurde pro Primerpaar eine Positiv-, eine Negativ- sowie eine Umschreibkontrolle mitgeführt (RT Umschreibung Polymerase negativ). In der Positivkontrolle wurden Klone von jeweils spezifischen PCR-Produkten eingesetzt. Als Ligation diente der p-drive (Fa. Quiagen, Hilden). Der Klon wurde mittels Elektroporation hergestellt. Das Programm des Thermocyclers (Abi Prism 7500, Fa. Applied Biosystems, Darmstadt) setzte sich folgendermaßen zusammen: Zyklus 1: (1x) Step 1: Zyklus 2: (45x) Denaturierung 95 °C 10 min. Amplifikation Step 1: 95 °C 15 sek. Step 2: 60°C 1 min. Zyklus 3: (1x) Schmelzpunktanalyse Step 1: 95 °C 15 sek. Step 2: 60 °C 1 min. Step 3: 95 °C 15 sek. 36 Eigene Untersuchungen Die Auswertung erfolgte mithilfe des Computerprogramms 7500 System SDS Software (Fa. Applied Biosystems, Darmstadt) anhand der Ct-Werte und Kontrolle der Schmelzkurve. Die spezifischen Schmelzpunkte für die untersuchten Erreger lagen bei: Enteroviren: 84.8-85,1 °C Adenoviren: 87,7-87,8 °C Influenza A: 83,5-83,8 °C Influenza B: 81,9-82,2 °C RSV: 79,5-81,0 °C Aktin (Referenzgen): 88,6 °C 3.1.5.4 Nested PCR Im Gegensatz zur qRT-PCR diente hier Ethidiumbromid als Detektionssystem. Die Spezifitätsprüfung erfolge über die Gelelektrophorese mit anschließender Sequenzierung. Primer: Um die Ziel-cDNA spezifisch nachweisen zu können, mussten auch hier die eingesetzten Primer komplementär zur cDNA-Vorlage sein. Die verwendeten Primer waren 16 (Picornaviren), 21 (Masern) und 28 Nukleotide (Pan Paramyxovirus) lang. Aufgrund der spezifischen GC-Gehalte lag die Annealing-Temperatur für Picornaviren bei 55 °C, für Masern bei 54 °C und für Pan Paramyxo bei 50 °C. Bei den verwendeten Primern handelte es sich um folgende: Picornaviren: OL-26 s 1st round (5´-GCACTTCTGTTTCCCC-3´) OL-27 as 1st round (5´-CGGACACCCAAAGTAG-3´) OL-26 seminested (5´-GCACTTCTGTTTCCC-3´) JAW-1 b as seminested (5´-CATTCAGGGGCCGGAGGA-3´) (JANG et al. 2006) 37 Eigene Untersuchungen Masern: MaN 1 (5´-ggTyCggATggTTCgAgAACA-3´) MaN 2 (5´-gRTTCATCAAggACTCAAgTg-3´) MaN 3 (5´-TgAAgTgCAAgACCCTgAggg-3´) MaN 4 (5´-TTC ATg CAg TCC AAg AgC agg-3´) (SANTIBANEZ et al. 1999) Pan paramyxo: PAR-F1 1st round (5´-GAAGGITATTGTCAIAARNTNTGGAC-3´) PAR-R 1st round (5´-GCTGAAGTTACIGGITCICCDATRTTNC-3´) PAR-R seminested (5´-GAAGGITATTGTCAIAARNTNTGGAC-3´) PAR-F2 seminested (5´-GTTGCTTCAATGGTTCARGGNGAYAA-3´) (TONG et al. 2008) PCR-Bedingungen: Die Vorbereitung des PCR-Ansatzes erfolgte ebenfalls unter der Sterilbank. Der verwendete Mastermix ist in Anhang 9.16 aufgelistet. Pro Primerpaar wurde eine Positiv- und eine Negativkontrolle mitgeführt. In der Positivkontrolle wurden Klone von jeweils spezifischen PCR-Produkten eingesetzt. Als Ligation diente der p-drive (Fa. Quiagen, Hilden). Der Klon wurde mittels Elektroporation hergestellt. Das Programm des Thermocyclers „Flex Cycler“ (Fa. Analytik Jena, Jena) setzte sich folgendermaßen zusammen: Zyklus 1: (1x) Step 1: Zyklus 2: (40x) Denaturierung 95 °C 5 min. Amplifikation Step 1: 95 °C 1 min. Step 2: 50, 54 oder 55 °C 40 sec. Step 3: 72 °C 1 min. Zyklus 3: (1x) Abschluss Step 1: 72 °C 4 min. Step 2: 4 °C unendlich 38 Eigene Untersuchungen Zur weiteren Spezifizierung wurde das PCR-Produkt für die Sequenzierung aufgearbeitet und analysiert (3130xl Genetic Analyzer, Fa. Life Technology, Darmstadt). Nach der Auftrennung des PCR Produkts mittels Elektrophoresekammer erfolgte das Herausschneiden und Weiterverarbeiten der spezifischen Banden unter UV-Licht (Millipore Ultrafree®-DA; Fa. Carl Roth GmBH, Karlsruhe). Im Anschluss an die Zentrifugation wurde das vom Gel getrennte PCR-Produkt durch eine Ethanolfällung nochmals aufgereinigt und unter Zugabe von 10 µl H2O konzentriert, die Proben einzeln auf ein 1 % Agarosegel aufgetragen und für fünf Minuten bei 90 Volt aufgetrennt (Elite 300 Plus; Fa. Schütt Labortechnik, Göttingen). Zur Konzentrationsbestimmung wurde eine spezifisch bekannte DNA Menge von 10, 30 und 50 ng eingesetzt. In einer speziellen Sequenzierungs-PCR lagerte sich das BigDye® Terminator v1.1 Cycle Sequencing Kit (Fa. Applied Biosystems, Darmstadt) an die einzelnen Nukleotide an. Nach einer erneuten Ethanolfällung und Trocknung war das Endprodukt bereit zur Sequenzierung. 3.1.6 Datenerfassung und -auswertung Für die mikroskopische Auswertung und die fotografische Dokumentation pathologischer Lungenveränderungen wurde das Lichtmikroskop Axioplan 2 Imaging (Fa. Zeiss, Oberkochen), die Digitalkamera AxioCam (Fa. Zeiss, Oberkochen) sowie das Computerprogramm AxioVision (Fa. Zeiss, Oberkochen) verwendet. Die Auswertung wurde an vorliegenden, archivierten Schnitten sowie nach Schnittherstellung von archivierten Paraffinblöcken und formalinfixiertem Gewebe durchgeführt. Die Erstellung der pathomorphologischen Diagnose erfolgte anhand der gängigen humanpathologischen Nomenklatur (BÖCKER 2008), modifiziert nach dem Lehrbuch „Pathologic basis of veterinary diseases“ (MCGAVIN u. ZACHARY 2007), wobei die Lungenalterationen nach Schweregrad, Dauer, Verteilung, Entzündungszelltyp und Art der Läsionen eingeteilt wurden. Die Unterscheidung zwischen interstitiellen Pneumonien und lediglich einer Ansammlung von minimalem Entzündungszellinfiltrat in den Alveolarsepten erfolgte histologisch über die Bestimmung der Septendicke. Eine Verbreiterung der Septen von 15 bis 30 µm (gemessen bei hundertfacher Vergrößerung) wurde als minimale Zellinfiltration 39 Eigene Untersuchungen beschrieben, wogegen eine Zunahme der Septendicke über 30 µm als interstitielle Pneumonie klassifiziert wurde. Die Graduierung der interstitiellen Pneumonien erfolgte histologisch mittels Photomikroskop. Anhand des in Tab. 3 abgebildeten Bewertungsschemas wurde pro Tier an zwei zufällig ausgewählten Bereichen bei hundertfacher Vergrößerung die Breite der Alveolarsepten gemessen und der Durchschnitt ermittelt. Tab. 3: Bewertungsschema zur semiquantitativen Beurteilung interstitieller Pneumonien. Grad Septenbreite geringgradig 30-50 µm gering- bis mittelgradig 50-100 µm mittelgradig 100-150 µm mittel- bis hochgradig 150-200 µm hochgradig über 200 µm Sekundäre Veränderungen, wie z. B. Emphyseme, Atelektasen und Verkalkungen, die nicht in direktem Kontakt zum Krankheitsgeschehen stehen, wurden unter dem Begriff „side effects“ zusammengefasst. Die gesammelten Daten wurden zusammen mit den Ergebnissen der ätiologischen Untersuchungen (Mikrobiologie, Parasitologie, Molekularbiologie) und tierspezifischen Informationen (Alter, Geschlecht, Vorbericht, Haltung) in Excel-Tabellen überführt und ausgewertet. Dies ermöglichte Aussagen über das Ausmaß verschiedener Lungenveränderungen in unterschiedlichen Tiergruppen sowie über eine Erregerbeteiligung. Um darzustellen, ob eine Geschlechts- und/oder Altersdisposition für pathologische Lungenveränderungen vorlag und um einen möglichen haltungsbedingten Einfluss zu untersuchen, wurden für einige Bereiche die Daten teils codiert, in Gruppen zusammengefasst und mithilfe der Filterfunktion sortiert. So sind die Weißbüschelaffen nach Herkunft (DPZ oder extern), Geschlecht (männlich, weiblich) und Alter unterteilt worden. Die einzelnen Altersstufen setzten sich wie folgt zusammen: Neugeborene Tiere waren 1 Tag bis 10 Wochen alt. Die nächste Altersgruppe bildeten die juvenilen Tiere von 10 Wochen 40 Eigene Untersuchungen bis 30 Monate. Adulte Weißbüschelaffen waren alle über 30 Monate alten Tiere. Grundsätzlich wurde das bei der Sektion entnommene Organspektrum histologisch ebenfalls untersucht, um einen Gesamteindruck über den Gesundheitsstatus der Tiere zu erlangen. Die Dokumentation dazu erfolgte in abteilungseigenen Untersuchungsprotokollen und wurde in dieser Arbeit nicht weiter aufgeführt. Die Ausnahme bildeten einzelne histologische Befunde, die bei der Auswertung ätiologischer Untersuchungsergebnisse von Relevanz waren. Immunhistochemische Untersuchungen wurden an paraffiniertem Lungengewebe ausgewählter Tiere durchgeführt. Sie dienten zum einen der Darstellung infizierter Zellen und zum anderen einer quantitativen Bestimmung der an der interstitiellen Pneumonie beteiligten B- und T-Lymphozyten. Neben der Tierkolonie des Deutschen Primatenzentrums wurden in diese Untersuchung 38 extern gehaltene Weißbüschelaffen des Institutes für Zoologie und Anthropologie der Universität Göttingen und weitere 34 Tiere eines Unternehmens aus der Pharmaindustrie einbezogen (Tab. 4). Die Haltungsbedingungen der Tiere unterschieden sich insoweit, als die des DPZ und aus der Industrie unter konstanten klimatischen Bedingungen in einer Indoor-Haltung untergebracht werden, während die Universitätstiere zwar auch in beheizten Räumen gehalten wurden, aber jederzeit Zugang zu einem Außenbereich hatten. Die Gruppe der Universitätstiere setzte sich aus 27 männlichen und elf weiblichen Weißbüschelaffen zusammen. Von sechs Tieren war das Alter nicht dokumentiert, 18 waren juvenil und 14 adult. Die 34 Tiere des Industrieunternehmens kamen zwischen 2004 und 2010 zur Sektion ins DPZ, deren Haltungsbedingungen waren mit denen des DPZ vergleichbar. Die Gruppe setzte sich aus 12 Weibchen und 36 Männchen zusammen. Zwei Weißbüschelaffen waren neugeboren, 20 juvenil und 12 adult. Bei einem Tier war das Alter nicht dokumentiert. 41 Eigene Untersuchungen Tab. 4: Übersicht über die externen Weißbüschelaffen. Tiere der Universität Göttingen Tiere der externen Labortiereinrichtung Männlich Weiblich Männlich Weiblich Neugeboren 0 0 1 1 Juvenil 13 5 13 7 Adult 8 6 7 4 Unbekannt 6 0 1 0 Gesamt 27 11 22 12 3.1.7 Statistik Die Grundlage der statistischen Auswertung bildeten tierspezifische Informationen zusammen mit den pathomorphologischen Diagnosen. Zu ermitteln ist der Einfluss des Alters, des Geschlechts oder der Haltungsbedingungen auf das Auftreten minimaler mononukleärer Infiltrate in den alveolären Septen und interstitiellen Pneumonien. Mittels Häufigkeitstabellen wurden Unterschiede zwischen den verschiedenen Tiergruppen dargestellt, welche mithilfe des Chi-Quadrat-Tests auf Signifikanz überprüft wurden. Die statistische Auswertung erfolgte mit Microsoft® Excel® 2010 und Statistica 10 (Fa. Statsoft GmbH, Hamburg). 42 Ergebnisse 4 Ergebnisse Der Ergebnisteil der vorliegenden Untersuchung gliedert sich in drei Abschnitte. Der erste Teil gibt einen Überblick über die Bestandsentwicklung der Weißbüschelaffen im DPZ und über die Tiere, die in der Auswertung berücksichtigt wurden. Der Schwerpunkt dieser Arbeit lag auf dem zweiten Abschnitt, welcher sich mit dem Auftreten von Lungenveränderungen und deren quantitativer und qualitativer Analyse in den verschiedenen Tiergruppen des DPZ (Alter/Geschlecht/Haltung) beschäftigt. Berücksichtigung fanden ebenfalls die molekularbiologischen und bakteriologischen Untersuchungsergebnisse. Im letzten Abschnitt wurden zwei externe Tiergruppen untersucht, die im Rahmen von wissenschaftlichen Untersuchungen am DPZ euthanasiert und seziert wurden. 4.1 Allgemeine Angaben zu den untersuchten Weißbüschelaffen des DPZ Entwicklung des Bestandes und Untersuchungsmaterial Die Weißbüschelaffenkolonie des DPZ wurde 1984 mit 20 Tieren gegründet. Die Bestandsgröße wuchs in den folgenden Jahren stark an und variierte im Untersuchungszeitraum von 1997 bis 2011 zwischen 368 und 687 Tieren (Abb.1). Die Gesamtzahl der ausgewerteten Tiere lag bei 639, wobei die Anzahl der Tiere, die in diese Studie einbezogen wurden, in den jeweiligen Jahren stark schwankte. So lag sie im Jahr 2001 bei 16 und 1999 bei 100 Tieren. Für diese Arbeit wurden nur Tiere berücksichtigt, von denen neben allgemeinen Daten wie Geschlecht, Alter, Haltung, Sektionsmaterial und Arbeitsgruppe bekannt war, dass sie zuvor nicht an Studien teilgenommen hatten, welche sich möglicherweise auf die Morphologie der Lunge hätten auswirken können. Somit lag die Zahl der sezierten Tiere über derjenigen, die in diese Auswertung eingeflossen ist. Für die Sektion lagen unterschiedliche Gründe vor. Die Auswertung der Anamnesebögen zeigte, dass die Euthanasie der Tiere entweder zur Organentnahme oder aus Tierschutzgründen bei progressiver Verschlechterung des Allgemeinbefindens geschah. Bei Weißbüschelaffen mit dokumentierten klinischen Symptomen im Vorbericht handelte es sich in der Regel um gastroenterale Symptome. Nur bei drei Weißbüschelaffen wurden anamnestisch respiratorische Probleme erwähnt, die bei zwei Tieren die Todesursache darstellten. 43 Ergebnisse Ein kleiner Teil der Tiere verstarb plötzlich, ohne dass Symptome beobachtet wurden. Abb. 1: Koloniegröße und Anzahl der untersuchten Tiere. 700 600 Anzahl der TIere 500 400 300 200 100 83 100 59 26 16 22 47 34 29 24 50 48 48 24 29 0 1997 1998 1999 2000 2001 2002 2003 2004 2005 2006 2007 2008 2009 2010 2011 Koloniegröße Jahr untersuchte Tiere Geschlechts und Altersverteilung Von den 639 Tieren, die in die Untersuchung einbezogen wurden, waren 317 männlich (49 %) und 321 weiblich (51 %). Bei einem Tier ging das Geschlecht nicht aus der Dokumentation hervor. Die unterschiedlichen Altersstufen aller verstorbenen und euthanasierten Tiere sind in Abb. 2 dargestellt. Die einzelnen Klassen setzten sich wie folgt zusammen: Tiere von 1 Tag bis 10 Wochen zählten zur Gruppe der Neugeborenen, welche 170 Weißbüschelaffen umfasste (27 %). In der Altersklasse der Juvenilen, im Alter von 10 Wochen bis 30 Monate, befanden sich 141 Tiere (22 %). Über 30 Monate waren 328 Individuen und bildeten die Altersstufe der Adulten (51 %). 44 Ergebnisse Abb. 2: Altersverteilung der untersuchten Tiere des DPZ. 350 328 300 Anzahl der Tiere 250 200 170 141 150 100 50 0 Neugeboren Juvenil Adult Alter 4.2 Überblick über die Lungenveränderungen im Tierbestand des DPZ Im Untersuchungszeitraum von 1997 bis 2011 wurden die Daten von 639 Weißbüschelaffen des DPZ in diese Untersuchung einbezogen. Mit 343 Tieren zeigte ca. die Hälfte (54 %) histopathologisch keine besonderen Befunde (o. b. B.) in der Lunge. Die übrigen Tiere zeigten unterschiedliche entzündliche Veränderungen und Pneumonieformen. In fünf Fällen wurden Lungentumoren nachgewiesen. Die Veränderungen wurden eingeteilt in: minimale interstitielle Entzündungsinfiltrate (n= 66) interstitielle Pneumonien (n= 205) alveoläre Herdpneumonien (n= 18) mit purulenten Bronchopneumonien, Lobärpneumonien und Alveolitiden bronchointerstitielle Pneumonien (Mischform) (n= 2) sonstige Pneumonien (n= 1: granulomatöse Pneumonie) Von allen untersuchten Lungenveränderungen waren die interstitiellen Pneumonien bei 205 Tieren (32 %) - am häufigsten vorhanden. Eine genaue Betrachtung der 45 Ergebnisse interstitiellen Pneumonien und der minimalen interstitiellen Entzündungsinfiltrate erfolgt in den nachfolgenden Abschnitten 4.2.1 und 4.2.2. Alveoläre Pneumonien (alv. Pn.) konnten bei insgesamt 18 Tieren (3 %) nachgewiesen werden. Diese setzen sich aus sechs Bronchopneumonien, neun Lobärpneumonien und drei Alveolitiden zusammen. Die Bronchopneumonien (Tab. 11, Anhang) traten bei vier Weibchen und zwei Männchen auf, wobei jeweils zwei neugeborene, juvenile und adulte Tiere betroffen waren. Aufgrund der Dominanz der neutrophilen Granulozyten, handelte es sich hierbei um purulente Bronchopneumonien (Abb. 3). Bei allen betroffenen Weißbüschelaffen trat eine hochgradige Ausprägung der Entzündung auf. Das Verteilungsmuster variierte von fokal bis diffus. Mit vier Tieren zeigte die Mehrzahl der Weißbüschelaffen einen akuten bis subakuten Verlauf. Ein Tier wies eine akute Entzündung und ein weiteres eine akute bis subakute Entzündung auf. 46 Ergebnisse Abb. 3: Lunge eines Weißbüschelaffen mit einer hochgradigen, akut bis subakuten, diffusen Bronchopneumonie (G-Nr.: 4643; Paraffinschnitt, H.E.; Messbalken 100 µm). Lobärpneumonien (Abb. 4 und Tab. 12, Anhang) zeigten sich bei vier Weibchen und drei Männchen. Darunter waren zwei neugeborene, zwei juvenile und drei adulte Tiere. Anhand der Entzündungszellen wurde zwischen purulenten (2 Tiere), fibrinöspurulenten (2 Tiere) und fibrinösen Lobärpneumonien (3 Tiere) unterschieden. Der Schweregrad variierte von geringgradig (1 Tier), über gering bis mittelgradig (1 Tier) und mittelgradig (3 Tiere) bis hochgradig (2 Tiere). Mutlifokal-konfluierende Entzündungen wurden am häufigsten diagnostiziert (4 Tiere). Multifokale Ausprägungen traten bei zwei Tieren und fokale Verteilungen bei einem Tier auf. Zwei weitere adulte Weibchen wiesen mittel- bis hochgradige Pneumonien auf und multifokal war die angrenzende Pleura betroffen. 47 Ergebnisse Abb. 4: Tier mit einer hochgradigen, akut bis subakuten, multifokal bis konfluierenden Lobärpneumonie (G-Nr.: 6382; Paraffinschnitt, H.E.; Messbalken 100 µm). 48 Ergebnisse Alveolitiden (Tab. 13, Anhang) stellten sich bei zwei Männchen und einem Weibchen dar. Zwei Tiere zeigten gering- und mittelgradige subakute Verläufe mit lymphoplasma-histiozytären Zellinfiltraten im Lungenparenchym. Ein männlicher Weißbüschelaffe wies eine geringgradige, akute Verlaufsform mit überwiegend neutrophilen Granulozyten auf. Bronchointerstitielle Lungenentzündungen (Tab. 14, Anhang) mit histopathologischen Eigenschaften von Bronchopneumonien und interstitiellen Pneumonien wurden bei einem adulten Männchen und einem juvenilen Weibchen diagnostiziert. Während beim männlichen Tier eine hochgradige subakute diffuse Entzündung vorlag, war beim Weibchen eine mittelgradige multifokale akute Pneumonie zu beobachten. Bei einem adulten Weibchen in Abb. 5 (Tab. 15, Anhang) ließ sich eine granulomatöse Lungenentzündung als Reaktion auf aspiriertes Fremdmaterial (Haar) darstellen. 49 Ergebnisse Abb. 5: Fremdkörpergranulom in der Lunge eines Weißbüschelaffen (G-Nr.: 6382; Paraffinschnitt, H.E.; Messbalken 10 µm). Fünf Weißbüschelaffen (Tab. 16, Anhang) wiesen Lungentumoren auf, wovon es sich um ein Fibrosarkom, ein malignes B-Zell Lymphom, ein T-zellreiches B-Zell Lymphom und ein nicht näher klassifiziertes Lymphom handelte. Abb. 6 stellt ein Lymphom in der Lunge dar, welches mithilfe immunhistochemischer Untersuchungen als B-Zelllymphom klassifiziert wurde. Bronchointerstitielle und granulomatöse Pneumonien sind zusammen mit den Tumoren in Abb. 7 unter der Gruppe „sonstige“ zusammengefasst (1 %). 50 Ergebnisse Abb. 6: Lunge eines Weißbüschelaffen mit einem malignen B-Zelllymphom (G-Nr.: 6758; Paraffinschnitt, H.E.; Messbalken 100 µm). 51 Ergebnisse Abb. 7: Verteilung der Lungenveränderungen. alv. Pn. 3% sonstige 1% interstitielle Pneumonie 32% o.b.B. 54% minimales Zellinfiltrat 10% Um spezielle Erregerstrukturen darstellen zu können, wurden in ausgesuchten Fällen Spezialfärbungen angefertigt. Hierbei handelte es sich um Pneumonien, bei denen Bakterien oder Pilze als auslösendes Agens vermutet wurden. Bei einem juvenilen, männlichen Weißbüschelaffen mit einer geringgradigen, akuten, multifokalen Alveolitis konnten Pilzhyphen mit der Versilberung nach Grocott nachgewiesen werden, zurückzuführen auf eine generalisierte Candidose, die ihren Ursprung vermutlich im Darm hatte und sich in Folge einer lang andauernden antibakteriellen Behandlung manifestierte. Bei den bronchointerstitiellen Lungenentzündungen, den Bronchopneumonien und den Lobärpneumonien dienten die Periodic-Acid-Schiff Reaktion und die GiemsaFärbung der Darstellung verschiedener Bakterien. Bordetella bronchiseptica, Streptokokken und Enterokokken konnten mithilfe dieser Methoden angefärbt und durch mikrobiologische Untersuchungen bestätigt werden. Ein weibliches Tier mit einer mittelgradigen, chronisch-aktiven, multifokalen Lobärpneumonie zeigte histologisch dichte Bakterienkolonien, die nicht kultivierbar waren. Aufgrund der Erregermorphologie wurde der Verdacht auf eine Aktinomykose oder Nokardiose geäußert. Leider war eine genauere Speziesbestimmung nicht möglich. 52 Ergebnisse 4.2.1 Untersuchung der minimalen Infiltrationen mit Entzündungszellen Bei 10 % der untersuchten Weißbüschelaffen (66 Tiere) konnte in den Alveolarsepten eine minimal erhöhte Infiltration mit vornehmlich mononukleären Entzündungszellen festgestellt werden. Bei diesen handelte es sich vor allem um Makrophagen, Lymphozyten und wenige Plasmazellen, die zu einer Verbreiterung der Septen (15-30 µm, gemessen bei hundertfacher Vergrößerung) führte. Anhand der Bestimmung der Entzündungszellen erfolgte eine semiquantitative Einteilung in sechs Stadien (Tab. 5). Fast alle Tiere (86 %) wiesen einen subakuten Verlauf auf. Weniger häufig wurden akute-subakute (fünf Tiere = 8 %) und akute Verläufe (vier Tiere = 6 %) beschrieben (Abb. 19, Anhang). Tab. 5: Charakterisierung der Chronizität anhand der Entzündungszellen. Chronizität dominierende Entzündungszellen akut neutrophile Granulozyten akut-subakut neutrophile Granulozyten, Makrophagen, Lymphozyten, Plasmazellen subakut Lymphozyten, Makrophagen, Plasmazellen subakut-chronisch Lymphozyten, Makrophagen, Fibroblasten chronisch Fibroblasen, Lymphozyten, Makrophagen chronisch-aktiv Fibroblasten, neutrophile Granulozyten, Makrophagen, Lymphozyten Mit 37 Tieren (56 %) zeigte über die Hälfte der Tiere mit milden entzündlichen Veränderungen des Interstitiums ein multifokales Verteilungsmuster. Am zweithäufigsten wurde bei insgesamt 16 Tieren (24 %) eine fokale Ausprägung festgestellt. In geringerem Maße konnten multifokal-konfluierende (sechs Tiere), oligofokale (fünf Tiere) und diffuse (zwei Tiere) Verteilungen der entzündlichen Infiltrate beschrieben werden (Abb. 20, Anhang). In Bezug auf das Geschlecht wurde bei den insgesamt 66 Weißbüschelaffen keine Prädisposition festgestellt. 34 weibliche (52 %) und 32 männliche Tiere (48 %) wiesen minimale Entzündungszellinfiltrate in den Lungen auf (Tab. 17, Anhang). 53 Ergebnisse Bei der Auswertung der Altersverteilung fiel auf, dass, wie in Abb. 8 dargestellt, 31 adulte Tiere (47 %) betroffen waren. In der Gruppe der Neugeborenen wiesen 20 Tiere (30 %) und bei den Juvenilen 15 Tiere (23 %) die beschriebenen Veränderungen auf. Da prozentual gesehen diese Verteilungen in gewissem Maße der Altersverteilung der Gesamtpopulation entsprachen (Abb. 8), wurde in Tab. 6 das Auftreten von minimalen Entzündungszellinfiltraten in Abhängigkeit zur Gesamtpopulation untersucht. Hierbei zeigte sich, dass nur minimale Abweichungen zwischen den einzelnen Altersklassen auftraten und mit 12 % die neugeborenen Tiere etwas häufiger betroffen waren als die juvenilen (11 %) oder die adulten (9 %). Eine Altersprädisposition ließ sich auch nicht durch den Chi-Quadrat-Test (p = 0,715) feststellten. Abb. 8 Altersverteilung der minimalen Infiltrationen mit Entzündungszellen. Neugeboren 30% Adult 47% Juvenil 23% 54 Ergebnisse Tab. 6: Altersabhängige Verteilung im Verhältnis zur Gesamtpopulation. kein Entzündungszellinfiltrat minimales Entzündungszellinfiltrat Gesamt Neugeboren 149 20 169 % 88 12 Juvenil 126 15 % 89 11 Adult 296 31 % 91 9 Gesamt 571 66 Zusammenfassend handelte es sich bei den 141 327 637 minimalen Infiltrationen im Lungeninterstitium bei der Mehrzahl der untersuchten Weißbüschelaffen um mononukleäre Zellen, wie sie bei einem subakuten Verlauf auftreten, bestehend aus Makrophagen, Lymphozyten und wenigen Plasmazellen mit multifokaler Verteilung, wie in Abb. 9 dargestellt. Abb. 10 dient als Vergleich mit einem lungengesunden Tier. Eine Alters- oder Geschlechtsprädisposition konnte nicht festgestellt werden. Hinweise auf Infektionserreger fanden Schnittpräparate nicht. 55 sich anhand der histologischen Ergebnisse Abb. 9: Lunge eines Weißbüschelaffen ohne besonderen Befund (G-Nr.: 8553; Paraffinschnitt, H.E., Messbalken 50 µm). 56 Ergebnisse Abb. 10: Lunge mit Infiltrationen mit Entzündungszellen in den Septen und einem geringgradigen alveolären Ödem. Erste Übergänge von minimalen Infiltraten zur interstitiellen Pneumonie (G-Nr.: 8000; Paraffinschnitt, H.E.; Messbalken 50 µm). 4.2.2 Untersuchung der interstitiellen Pneumonien Mit 205 Fällen bei 639 untersuchten Weißbüschelaffen (32 %) handelte es sich bei der interstitiellen Pneumonie um die am häufigste diagnostizierte Lungenveränderung. Zur Erstellung der morphologischen Diagnose wurden die gleichen Bewertungskriterien wie im vorherigen Kapitel 4.2.1 angewandt, somit erfolgte die Unterteilung in die sechs Chronizitätsstadien wie in Tab. 5. Anhand der Auswertung der interstitiellen Entzündungszellen stellte sich heraus, dass über die Hälfte der Tiere (59 %) einen subakuten Krankheitsverlauf aufwiesen. Als Besonderheit ist anzumerken, dass aus dieser Gruppe bei 2/3 der Tiere (66 %) neben den für dieses Stadium charakteristischen Zellen wie Lymphozyten, 57 Ergebnisse Makrophagen und vereinzelt Plasmazellen in geringem Maße auch neutrophile Granulozyten nachgewiesen wurden. Die übrigen Entzündungsstadien traten in geringerem Umfang auf, so waren, wie in Abb. 11 dargestellt, die subakutchronischen Entzündungen mit neun Fällen (4 %) am wenigsten und die chronischaktiven Entzündungen mit 24 Fällen (12 %) am häufigsten vertreten. Abb. 11: Chronizität der interstitiellen Pneumonien. chronisch-aktiv 12% akut 9% akut-subakut 7% chronisch 9% subakutchronisch 4% subakut 59% Die Graduierung der interstitiellen Pneumonien erfolgte histologisch anhand des in Tab. 3 abgebildeten Bewertungsschemas. Mit 131 Fällen bei 205 Tieren (64 %) mit interstitiellen Pneumonien sind die geringgradigen Ausprägungen dieser Entzündung am häufigsten vertreten, wogegen nur 26 Weißbüschelaffen (13 %) gering- bis mittelgradige Entzündungen zeigten. Am zweithäufigsten wurden bei 41 untersuchten Weißbüschelaffen (20 %) mittelgradige interstitielle Pneumonien diagnostiziert. Mittel- bis hochgradige und hochgradige Entzündungen kamen mit 5 (2 %) und 2 Fällen (1 %) sehr selten vor (Abb. 21, Anhang). Wie in Abb. 12 dargestellt wurden die Ausprägungen in fünf verschiedene Kategorien unterteilt. Bei 38 Tieren (19 %) war herdförmig nur ein kleiner Lungenbereich betroffen (fokale Ausprägung). Eine oligofokale Verteilung, d. h. 1-3 betroffene 58 Ergebnisse Bereiche, konnte bei 10 Weißbüschelaffen (5 %) diagnostiziert werden. Mit 63 Tieren (31 %) und 89 Tieren (43 %) traten die multifokalen, mit mehr als 3 Entzündungsherden, und die multifokal-konfluierenden interstitiellen Pneumonien am häufigsten auf. Mit diffusem Muster wurden Entzündungen beschrieben, bei denen sich die Entzündungszellinfiltrate gleichmäßig über die gesamte Lunge verteilten, ohne dass noch unveränderte Bereiche zu erkennen waren. Diese Form trat bei lediglich 5 Tieren (2 %) auf. Hinweise auf Infektionserreger fanden sich anhand der histologischen Schnittpräparate nicht. Abb. 12: Verteilung der interstitiellen Pneumonien. 100 89 90 80 Anzahl der Tiere 70 63 60 50 40 38 30 20 10 10 5 0 fokal oligofokal multifokal multifokalkonfluierend diffus Verteilungsmuster Um Aussagen über eine mögliche Geschlechtsprädisposition zu treffen, wurde das Auftreten interstitieller Pneumonien bei männlichen und weiblichen Tieren getrennt voneinander untersucht. Hierbei zeigte sich, dass insgesamt 104 weibliche (51 %) und 101 männliche Tiere (49 %) erkrankten. Dieses Verhältnis entsprach prozentual dem der Gesamtpopulation (Tab. 18, Anhang). 59 Ergebnisse Bei der Untersuchung der drei Altersklassen wurden 51 interstitielle Pneumonien bei neugeborenen, 60 bei juvenilen und 94 bei adulten Weißbüschelaffen diagnostiziert. Um eine eventuelle Häufung in den verschiedenen Altersklassen zu prüfen, wurde gruppenintern die Anzahl der interstitiellen Lungenentzündungen in Bezug zur restlichen Altersgruppe gesetzt. Wie in Tab. 7 dargestellt waren mit 42 % besonders juvenile Tiere betroffen, während Neugeborene und Adulte nur zu 30 % bzw. 28 % betroffen waren. Signifikanzen konnten ebenfalls statistisch durch den Shi-QuadratTest belegt werden. Da der p-Wert unter 0,05 (p = 0,0126) liegt besteht ein Zusammenhang zwischen dem Alter der Tiere und der Wahrscheinlichkeit an interstitiellen Pneumonien zu erkranken (Tab. 19, Anhang) Tab. 7: Altersverteilung der Tiere mit interstitieller Pneumonie im Verhältnis zur Gesamtgruppe. Tiere ohne interstitielle Pneumonie Tiere mit interstitieller Pneumonie Gesamt Neugeboren 118 51 30 % 169 Juvenil 81 60 42 % 141 Adult 233 94 28 % 327 Gesamt 432 205 637 Da die Grundlage dieser Dissertation die Daten von 1997 bis 2011 bildeten, die Weißbüschelaffen aber im April 2004 in einer neuen Tiereinheit untergebracht worden sind, wurde dieser Stichtag verwendet, um mögliche Unterschiede bei den Pneumonien zwischen alter und neuer Haltung zu beurteilen. Tiere nach April 2004, die zeitweise in der einen oder anderen Einheit untergebracht wurden, sind in dieser Erhebung nicht mit berücksichtigt. Daher liegt, wie in Tab. 8 dargestellt, die Gesamtzahl für diese Untersuchung mit 585 Weißbüschelaffen unter der in Kapitel 4.1 erwähnten Zahl von 639. „Haltung 1“ bezeichnet das alte Tierhaus. Hier flossen Daten von 476 Primaten in die Auswertung ein, wohingegen es in „Haltung 2“, also nach April 2004, 109 Tiere waren. Auffällig ist, dass die Zahl der Weißbüschelaffen 60 Ergebnisse mit interstitiellen Pneumonien im Untersuchungszeitraum von 29 %, in der alten Haltung, um 14 Prozentpunkte auf 43 % in der neuen Haltung angestiegen ist (Tab. 8). Damit geht entsprechend die Abnahme mit der Zahl der Tiere einher, welche ohne besondere Befunde in der Lunge waren (Tab. 9). Durch die statistische Untersuchung mittels Chi-Quadrat-Test konnten ebenfalls Signifikanzen festgestellt werden, da der p-Wert mit 0,006 unter der Grenze von 0,05 lag. Tab. 8: Auftreten von interstitiellen Pneumonien in Abhängigkeit zur Haltung. Haltung Tiere ohne interstitielle Pneumonie Tiere mit interstitieller Pneumonie Gesamt 1 336 140 476 % 71 29 2 62 47 % 57 43 Gesamt 394 187 109 585 Tab. 9: Auftreten von lungengesunden Tieren in Abhängigkeit zur Haltung. Haltung Tiere mit Lungenbefund Tiere ohne Lungenbefund Gesamt 1 213 263 476 % 45 55 2 60 49 % 55 45 Gesamt 273 312 109 585 Zusammenfassend ist festzuhalten, dass es sich bei den Lungenveränderungen im Bestand der Weißbüschelaffenkolonie des DPZ vor allem um Tiere mit geringgradigen, subakuten, multifokal-konfluierenden interstitiellen Pneumonien handelte, welche neben dem charakteristischen Zellinfiltrat eines subakuten 61 Ergebnisse zeitlichen Verlaufs ebenfalls neutrophile Granulozyten aufwiesen (Abb. 13). Signifikanzen konnten für kein Geschlecht festgestellt werden, wohingegen in der Altersgruppe der juvenilen Tiere ein gehäuftes Auftreten vorlag. Ebenso ließ sich ein Anstieg dieser Form der Lungenentzündung zwischen altem und neuem Tierhaus erkennen. Abb. 13: Lunge mit einer geringgradigen, subakuten, multifokal-konfluierenden interstitiellen Pneumonie. Im oberen Bildrand zeigt sich noch unverändertes Lungengewebe (G-Nr.: 8136; Paraffinschnitt, H. E.; Messbalken 50 µm). Bakteriologische Untersuchungen Bakteriologische Ergebnisse der Lunge lagen für 98 Tiere vor, wobei, wie in Tab. 10 dargestellt, 29 Weißbüschelaffen einen positiven Befund aufwiesen. Von diesen wiederum zeigte die Lunge von zehn Tieren histologisch keine Veränderungen, 62 Ergebnisse sodass hier von einer septikämischen Erregerstreuung oder einer Kontamination bei der Probenaufbereitung ausgegangen werden muss. Bei zehn weiteren Weißbüschelaffen wurden interstitielle Pneumonien diagnostiziert. Bei fünf Tieren konnte in der Lunge Escherichia coli festgestellt werden, der bei allen Weißbüschelaffen ebenfalls im Darmtrakt nachgewiesen wurde. Eine Lunge war geringgradig positiv für Streptococcus sp., eine für Erysipelothrix rhusiopathiae, eine weitere für Klebsiella pneumoniae und eine andere wies eine Mischinfektion, bestehend aus Klebsiella pneumoniae und Pseudomonas aeruginosa, auf. Bei neun alveolären Pneumonien konnten Escherichia coli, Enterococcus faecalis, Bordetella bronchiseptica, Klebsiella pneumoniae, Klebsiella ozeaneae und geringgradig Streptococcus sp. isoliert werden. Der Weißbüschelaffe mit dem Lymphom in der Lunge wies ebenfalls in verschiedenen Organen gering- bis hochgradig hämolysierende Escherichia coli auf, wofür hierbei vermutlich ein septisches Geschehen in Betracht kommt. Tab. 10: Übersicht über die Tiere mit positivem Bakteriologiebefund der Lunge. Anzahl der Tiere ohne besonderen Befund interstitielle Pneumonie alveoläre Pneumonie Tumor 10 9 9 1 Molekularbiologische Untersuchungen Mittels Real-Time PCR wurden 22 ausgewählte Lungengewebsproben auf Influenza A/B, Adenoviren, Enteroviren und das Respiratorische Synzytialvirus A/B, untersucht. Das gleiche Gewebe wurde auf Picornaviren, Paramyxoviren und Masernviren mithilfe einer nested PCR getestet. Während die Real-Time PCR keine Viren nachweisen konnte, verlief nur der Maserntest per nested PCR bei zwei Tieren positiv. Hierbei handelte es sich um weibliche, adulte Weißbüschelaffen (Tab. 20, Anhang). Bei dem jüngeren Tier (Nr.: 8506) blieb die Lunge ohne besonderen Befund. Bei dem älteren Tier (Nr.: 8524) fanden sich eine Alveolarhistiozytose und eine Leukozytostase der Alveolarsepten. Weitere Hauptbefunde konnten bei beiden 63 Ergebnisse Tieren im Darmtrakt erhoben werden. Der Dünndarm von dem jüngeren Weißbüschelaffen wies Veränderungen in Form einer hochgradigen chronischen, lymphozytären Entzündung auf, und der Dickdarm zeigte eine mittelgradige lymphozytäre Kolitis. Bei dem älteren Tier mit positivem Masernbefund stellte sich im Duodenum eine chronisch-aktive erosive Entzündung dar. Immunhistochemische Untersuchung auf B-Lymphozyten, T-Lymphozyten und Makrophagen Um festzustellen, ob die mononukleären Infiltrate in den Alveolarsepten bei den Tieren mit interstitieller Pneumonie vorwiegend aus B- oder T-Lymphozyten bestehen, wurden von der Lunge von vier Tieren immunhistochemische Untersuchungen angefertigt. Ebenfalls wurden die Makrophagen markiert. Bei diesen Tieren handelte es sich um zwei Weibchen und zwei Männchen aus der am häufigsten vertretenen Gruppe der juvenilen Weißbüschelaffen mit gering- und mittelgradigen, subakuten, multifokal-konfluierenden interstitiellen Pneumonien. Auffällig war, dass unabhängig vom Schweregrad kaum B-Lymphozyten vorhanden waren, und es sich bei dem mononukleären Infiltrat in den Septen fast ausschließlich um T-Lymphozyten handelte (Abb. 14 und Tab. 22, Anhang). Die Anzahl der Makrophagen variierte deutlich. Mit 602 Zellen wurden bei dem Tier mit der mittelgradigen Pneumonie (Nr. 7824) die wenigsten und mit 1192 bei dem Tier mit der geringgradigen Pneumonie (Nr. 7681) gemessen. 64 die meisten Entzündungszellen Ergebnisse Abb. 14: Immunhistochemische Untersuchung mit anti-CD3 Antikörpern bei einem Tier mit einer mittelgradigen, subakuten, multifokalen interstitiellen Pneumonie (G-Nr.: 7789; Paraffinschnitt; Messbalken 50 µm). Side effects Zu den regelmäßig festgestellten Nebenbefunden, die in den Lungen der untersuchten Tiere auftraten, gehörten Verkalkungen und pleurale Bindegewebsproliferationen, Pigmentspeicherungen, alveoläre und interstitielle Ödeme, Emphyseme, Atelektasen und Hyperämien. Regelmäßig zeigten sich zufällig im Lungenparenchym verteilte Verkalkungen, wobei diese sowohl in den alveolären Septen als auch subpleural auftraten (Abb. 15). In der überwiegenden Anzahl der Fälle handelte es sich um geringgradige, runde bis ovale Verkalkungen. Der Durchmesser variierte von mehreren Mikro- bis Millimeter großen Kalkablagerungen, welche teilweise schon makroskopisch diagnostizierbar waren. 65 Ergebnisse Bei stichprobenartigen Untersuchungen betroffener Tiere mit geringgradigen Lungenverkalkungen konnten histologisch in den übrigen Organen keine weiteren Kalkablagerungen festgestellt werden. In einem Fall mit hochgradiger, multifokaler interstitieller Lungenverkalkung stand diese Veränderung im Zusammenhang mit einer fortgeschrittenen Nephropathie, die zu einem Ungleichgewicht des KalziumPhosphat-Haushaltes und damit zu einer metastatischen Verkalkung geführt hatte. Abb. 15: Lunge mit multifokalen Kalkablagerungen im Lungenparenchym bei einem Tier mit Niereninsuffizienz (G-Nr.: 8596; Paraffinschnitt, von Kossa Färbung; Messbalken 10 µm). Regelmäßig traten ebenfalls geringgradige, oligofokale, pleurale und subpleurale Gewebsproliferationen bei Tieren mit und ohne besonderen Befund in der Lunge auf. Immunhistochemische Markierungen mit anti-Vimentin- und anti-smooth muscle 66 Ergebnisse actin-Antikörpern sowie Trichromfärbungen nach Masson-Goldner zeigten, dass es sich hierbei um Bindegewebe handelte. Selten Formalinpigment, häufiger Kohlenstoffpigment oder Hämosiderin konnte histologisch nachgewiesen werden. Geringgradige Hämosiderinablagerungen, gelegentlich in Verbindungen mit Hyperämien und Blutungen, fanden sich vor allem in Alveolarmakrophagen im Sinne von Siderophagen. Am häufigsten stellten sich peribronchial und in den tracheobronchialen Lymphknoten kleine schwarze Herde, bestehend aus Kohlenstoffpigment (Anthrakose) dar. Dieses war in der Regel von Makrophagen phagozytiert oder lag reaktionslos im Gewebe (Abb. 16). Abb. 16: Lunge eines Weißbüschelaffen mit einer peribronchialen und perivaskulären Anthrakose (GNr.: 4586; Paraffinschnitt, H. E.; Messbalken 20 µm). 67 Ergebnisse Vor allem alveoläre Ödeme, sehr vereinzelt auch interstitielle Ödeme und geringgradige Emphyseme im Randbereich, fanden sich bei der überwiegenden Anzahl der Tiere. Während diese Veränderungen Nebenbefunde von Lungenentzündungen darstellen können, handelt es sich bei den Tieren ohne weitere histologische Lungenbefunde vermutlich um agonale Veränderungen. Die in großem Maße festgestellten Atelektasen fanden sich unabhängig vom Gesundheitszustand und stellten höchstwahrscheinlich präparationsbedingte Artefakte dar. 4.3 Überblick über Lungenveränderungen bei externen Tieren Neben den Weißbüschelaffen aus der eigenen Kolonie wurden ebenfalls zwei externe Weißbüschelaffengruppen untersucht, die aus Gründen des Bestandsmanagements und Tierschutzes im DPZ seziert worden sind. Über die Auswertung dieser Tiere sollte ermittelt werden, ob die Lungenveränderungen im DPZ Tierbestand repräsentativ für die Spezies sind oder ob es sich um bestandsabhängige Kooperationspartners Probleme aus der handelte. Dies Pharmaindustrie waren und 38 34 Tiere teils frei eines lebende Weißbüschelaffen des Institutes für Zoologie und Anthropologie der Georg-August Universität Göttingen. Tiere des Industriepartners Die 34 Tiere aus einer externen Labortierhaltung kamen zwischen 2004 und 2010 zur Sektion ins DPZ. Wie Abb. 17 zeigt, wies die Mehrzahl der Tiere Lungenveränderungen (65 %) auf. Mit 18 Weißbüschelaffen (insgesamt 53 %) handelte es sich dabei zum großen Teil um interstitielle Pneumonien. Nur vier Tiere (12 %) wiesen in den alveolären Septen minimale Infiltrate mit Entzündungszellen auf. 68 Ergebnisse Abb. 17: Verteilung der Lungenveränderungen o.b.B. 35% interstitielle Pneumonie 53% minimales Zellinfiltrat 12% Bei den vier Tieren mit minimalem Zellinfiltrat in den Septen handelte es sich um männliche Weißbüschelaffen, von denen zwei juvenil und eines adult ist. Bei einem weiteren wurde das Alter nicht dokumentiert. Ein Individuum zeigte einen akuten bis subakuten Krankheitsverlauf, wohingegen bei den drei anderen subakute Prozesse diagnostiziert wurden. Das Verteilungsmuster der subakuten Veränderungen wies jeweils eine fokale, eine multifokale und eine multifokal-konfluierende Ausprägung auf. Der akute bis subakute Prozess war nur fokal ausgebildet, und das histologische Bild wurde von neutrophilen Granulozyten, geringen Mengen an Lymphozyten und Makrophagen bestimmt. Dagegen dominierten bei dem subakuten Geschehen Lymphozyten und Makrophagen, während neutrophile Granulozyten nur vereinzelt auftraten. Die 18 Tiere mit interstitiellen Pneumonien setzten sich aus sieben weiblichen und elf männlichen Weißbüschelaffen zusammen, wobei 2 neugeboren, 10 juvenil und 6 adult waren. Statistische Untersuchungen mit einem Shi-Quadrat-Test konnten weder für das Geschlecht (p = 0,642) noch für eine Altersklasse (p = 0,4) Signifikanzen feststellen, da bei beiden Analysen der p-Wert über 0,05 lag (Tab. 23 und 24, Anhang). Mit 66 % (12 Tiere) dominierten geringgradige Pneumonien. Nur drei Weißbüschelaffen (17 %) zeigten gering- bis mittelgradige Veränderungen. Drei weitere zeigten mittelgradige Pneumonien. 69 Bei der Auswertung der Ergebnisse Entzündungszellen stellte sich heraus, dass zwei Tiere akute bis subakute (11 %), 14 Tiere subakute (78 %) und zwei weitere chronische Entzündungen (11 %) aufwiesen. Charakteristische Entzündungszellen für subakute Pneumonien waren Lymphozyten, Makrophagen und wenige Plasmazellen. Bei 10 der 14 Tiere konnten neben den beschriebenen Zellen ebenfalls in geringem Maße neutrophile Granulozyten nachgewiesen werden. Das Verteilungsmuster erstreckte sich bei 13 Tieren auf multifokale Bereiche. Bei vier Tieren lag eine fokale Entzündung vor und nur bei einem Tier ein diffuses Muster. Somit waren insgesamt geringgradige, subakute, multifokale interstitielle Pneumonien am häufigsten ausgeprägt mit lympho-plasmahistiozytärem Infiltrat und vereinzelt neutrophilen Granulozyten. Histologisch und mikrobiologisch ließen sich im Lungengewebe keine Infektionserreger nachweisen. Tiere der Universität Göttingen Insgesamt 38 Weißbüschelaffen des Institutes für Zoologie und Anthropologie wurden im DPZ obduziert. Mit 33 Tieren (87 %) blieb die Mehrzahl ohne besonderen Lungenbefund. Nur drei Weißbüschelaffen (8 %) zeigten minimales Zellinfiltrat, ein Tier eine alveoläre Pneumonie und eines eine interstitielle Entzündung. Bei den drei Weißbüschelaffen mit minimalen Veränderungen handelte es sich um zwei juvenile männliche und ein juveniles weibliches Tier, bei denen die Entzündungszellinfiltrate eine fokal bis multifokal-konfluierende Verteilung aufwiesen. Bei einem Tier wurde anhand der überwiegend neutrophilen Entzündungsreaktion die Entzündung als akut eingestuft. Bei zwei weiteren Tieren waren auch Makrophagen und Lymphozyten zu sehen, daher wurde der zeitliche Verlauf als akut bis subakut klassifiziert. Bei dem Weißbüschelaffen mit der hochgradigen, fokalen fibrinopurulenten Pneumonie handelt es sich um ein juveniles Weibchen (Abb. 18). Auch wenn mikrobiologisch in der Lunge bei diesem Tier kein spezifischer Erregernachweis geführt werden konnte, wiesen die nekrotisierenden Entzündungen in Leber, Milz und Lunge auf das Vorliegen einer infektiösen Allgemeinerkrankung hin. Der Antigennachweis, zusammen mit dem morphologischen Bild, sprach am ehesten für das Vorliegen einer akuten Tularämie. Bei den anderen Tieren waren keine spezifischen Pathogene in den Lungen nachweisbar. 70 Ergebnisse Abb. 18: Hochgradige, akute, fokale, fibrinopurulente Pneumonie bei einem Tier (G-Nr.: 7037) der Universität Göttingen (Paraffinschnitt, H. E.; Messbalken 20 µm). Fasst man diese beiden Tiergruppen zusammen, so konnten in der Labortierhaltung des industriellen Kooperationspartners ähnliche Ausprägungsmuster einer interstitiellen Pneumonie wie am DPZ festgestellt werden, während die Tiere der Universität Göttingen, die auch „outdoor“ Veränderungen zeigten. 71 – Möglichkeiten hatten, kaum Diskussion 5 Diskussion Seit der Gründung des Deutschen Primatenzentrums (DPZ) im Jahre 1977 stellt die Krallenaffenkolonie mit 462 Tieren (Stand 31.12.2011) einen erheblichen Teil der insgesamt 1283 am DPZ gehaltenen nicht humanen Primaten dar. Dementsprechend liegt gerade für diese Primatenspezies eine bedeutende Menge an asserviertem Tiermaterial vor. In dieser Arbeit wurden die Sektionsberichte und das formalinfixierte Lungengewebe der Jahre 1997 bis 2011 retrospektiv aufgearbeitet und insgesamt 639 DPZ eigene Weißbüschelaffen als auch 72 Tiere aus externen Haltungen untersucht. Neben der retrospektiven histologischen Aufarbeitung, die den Schwerpunkt dieser Arbeit bildet, wurden ebenfalls mithilfe molekularbiologischer Methoden (Echtzeit-PCR und Nested-PCR) 22 Lungengewebsproben auf verschiedene virale Erreger getestet. Anhand der erhobenen Daten sollte untersucht werden, welche Lungenveränderungen bei den Tieren auftreten, ob sich Erreger aus dem Lungengewebe isolieren lassen und ob eine Häufung in verschiedenen Tiergruppen zu erkennen ist. Am Deutschen Primatenzentrum wiesen im genannten Untersuchungszeitraum 226 von 639 Weißbüschelaffen Pneumonien auf. Wie in Abb. 7 dargestellt, liegt die Prävalenz für diese Tiere bei 35 %. Diese ist deutlich höher als bei vergleichbaren Studien anderer Autoren. In einer älteren Veröffentlichung von 1983 untersuchten CHALMERS et al. (1983) das Auftreten von Lungenentzündungen zum einen bei in Gefangenschaft aufgewachsenen Weißbüschelaffen und zum anderen bei wild lebenden Tieren. Von 265 untersuchten Zuchttieren wiesen 71 Weißbüschelaffen (27 %) Pneumonien auf, aber nur 3 von 70 bei den wild lebenden Tieren (4 %). In den folgenden Jahren wurden zwei weitere Studien veröffentlicht, die sich ebenfalls mit spontanen pathologischen Veränderungen bei Weißbüschelaffen befassten. So prüften DINIZ et al. (1994) in einem Zeitraum von 20 Jahren bei 265 Tieren das Auftreten von Krankheiten in einer Callithrix jacchus-Kolonie im Zoo von Sao Paulo und stellten fest, dass nach Erkrankungen des Digestionstraktes Pneumonien mit 16 % am zweit häufigsten auftraten. In der neuesten Erhebung von 2009 verglichen DAVID et al. (2009) die pathologischen Untersuchungsbefunde von 129 sezierten Weißbüschelaffen. Die Tiere nahmen zu Lebzeiten - wie in dieser Dissertation - an 72 Diskussion keinen Experimenten teil. Insgesamt wurden nur bei neun Weißbüschelaffen (7 %) Pneumonien diagnostiziert. Von den 34 Tieren der externen Labortiereinrichtung, die unter den gleichen IndoorBedingungen gehalten wurden wie die Tiere des DPZ, wiesen 18 Weißbüschelaffen Pneumonien auf. Hier lag mit 53 % eine noch größere Prävalenz vor. Bei den Tieren der Universität mit Zugang zu einem Außenbereich wiesen nur 2 von 38 Tieren Pneumonien auf. Diese Ergebnisse zeigen, dass die Prävalenzen für Pneumonien in den verschiedenen Kolonien stark variieren und die des DPZ mit 37 % relativ hoch sind. Die deutlichen Abweichungen in Bezug auf Lungenentzündungen könnten zum einen in Differenzen in der Auswertung und Charakterisierung von Lungenveränderung und zum anderen in unterschiedlichen Haltungsbedingungen begründet sein. Die Keimbelastung, der die Weißbüschelaffen in Labortiereinrichtungen ausgesetzt sind, ist von einer Vielzahl von Faktoren abhängig. Um Krankheitsausbrüche zu verhindern, muss der Erregereintrag in die Tierhaltung so gering wie möglich gehalten und ein guter Gesundheitszustand der Tiere gewährleistet werden. Daher ist es wichtig, nur gesunden Personen nach Kleidungswechsel und bei Verwendung eines Mundschutzes Zutritt zu den Tierräumen zu gestatten. Neu zugehende Tiere sollten eine Quarantäne durchlaufen und alle Tiere sind engmaschig tierärztlich zu überwachen. Ebenso sind die klimatischen Anforderungen für diese tropische Affenart strikt einzuhalten. Um Immunsuppressionen vorzubeugen, sollten die Tiere geringstmöglichem Stress ausgesetzt, menschliche Eingriffe auf ein Minimum beschränkt und bei der Gruppenzusammensetzung der Tiere auf deren soziale Verträglichkeit geachtet werden. Von einigen Fallbeschreibungen abgesehen sind quantitative Erhebungen über das Auftreten verschiedener Pneumonieformen bei Weißbüschelaffen nicht dokumentiert. Die vorliegende Studie ergab, bei hauseigenen Tieren mit Lungenveränderungen am häufigsten interstitielle geringgradigen, Pneumonien subakuten, (32 % der multifokal-konfluierenden untersuchten Tiere ) Entzündungen. mit Deutlich weniger häufig traten minimale Entzündungszellinfiltrate in den Septen (10 % der untersuchten Tiere) auf. Die Pathogenese der interstitiellen Pneumonien ist komplex 73 Diskussion und kann auf einer Schädigung des Alveolarepithels (Typ 1- und Typ 2Pneumozyten), auf einer hämatogenen Schädigung des alveolären Kapillarendothels oder der alveolären Basalmembran beruhen. Somit kommen ätiologisch neben pneumo- bzw. endotheliothropen Viren vor allem bakterielle Toxine und Umwelteinflüsse, wie z. B. die Inhalation bestimmter Gase oder Schwebstoffe, in Betracht (MCGAVIN u. ZACHARY 2009). Nur bei wenigen Tieren konnten die Ursachen der interstitiellen Pneumonie festgestellt werden. Die verstorbenen Weißbüschelaffen mit interstitieller Pneumonie, die mikrobiologisch untersucht wurden, wiesen mit Streptococcus sp., Klebsiella pneumoniae, Escherichia coli, Pseudomonas aeruginosa und Erysipelothrix rhusiopathiae unterschiedliche Erreger von variabler Pathogenität auf. Hinsichtlich des Nachweises der Streptokokken sind beim Menschen und nicht humanen Primaten Streptococcus pneumoniae als obligat pathogener Keim bekannt (ABEE et al. 2012). Für den Weißbüschelaffen liegen keine Fallbeschreibungen mit diesem Infektionserreger vor. Bei anderen Neuweltaffen sind zwar keine interstitiellen Lungenentzündungen, aber schwere, letale Bronchopneumonien dokumentiert (BERENDT et al. 1975). Auch andere durch Streptokokken verursache Erkrankungen fanden sich bei diesem Tier nicht, weshalb hier von einer Kontamination ausgegangen wurde. CHALMERS et. al. (1983) untersuchten beim Weißbüschelaffen durch Klebsiella sp. verursachte Pneumonien, und stellten ebenfalls Bronchopneumonien und keine interstitiellen Lungenentzündungen fest. Diese zur Familie der Enterobacteriaceae gehörenden Stäbchenbakterien verursachen ebenfalls Enteritiden und konnten bei den zwei betroffenen Tieren aus dem entzündlich veränderten Darm isoliert werden. Insoweit handelte es sich vermutlich um eine septikämische Erregerstreuung. Fünf der sieben Tiere mit einem positiven E. coli Befund in der Lunge wiesen diesen Infektionserreger auch im Darm auf. Hier dürfte ebenfalls eine septikämische Ausbreitung vorliegen. Zwei der Tiere befanden sich zum Untersuchungszeitpunkt im Stadium fortgeschrittener Autolyse, weshalb in diesen beiden Fällen ein postmortaler Überwuchs nicht auszuschließen ist. Ein septikämisches Krankheitsgeschehen, vermutlich von der Gallenblase oder der Leber ausgehend, war auch beim Weißbüschelaffen mit positivem Pseudomonadenbefund zu vermuten. Bei einem 74 Diskussion weiteren Tier konnte Erysipelothrix rhusiopathiae in allen Organen nachgewiesen werden. Vermutlich war dies ebenfalls Ausdruck einer septikämischen Verbreitung, da ein entsprechendes pathomorphologisches Korrelat in der Lunge nicht festzustellen war. Um eine virale Genese für das gehäufte Auftreten von interstitiellen Pneumonien abzuklären, wurden in ausgewählten Fällen molekularbiologische Analysen durchgeführt. Bei den 22 Tieren, die mittels Real-Time und nested PCR auf Influenza A/B, Adenoviren, Respiratorisches Synzytialvirus A/B, Enterovirus, Picornaviren, Pan Paramyxoviren und Masernvirus getestet wurden, handelte es sich um eine heterogene Tiergruppe, bestehend aus männlichen und weiblichen Weißbüschelaffen, die sich aus neugeborenen, juvenilen und adulten Krallenaffen zusammensetzte. Im Lungengewebe von zwei dieser Weißbüschelaffen konnten Masernviren nachgewiesen werden (Tab. 21, Anhang). Infektionen mit dem Masernvirus sind für verschiedene Altweltaffen, wie Klammeraffen (Ateles spp.), Zwergmeerkatzen (Cercopithecus talapoin), Schimpansen, Rhesus- und Javaneraffen beschrieben (VAN BINNENDIJK et al. 1995; CHOI et al. 1999; EL MUBARAK et al. 2007). Bei den Neuweltaffen liegen Berichte für östliche GraukehlNachtaffen, Rotbauch- und Schnurrbarttamarine (Saguinus mystax) aber auch für Weißbüschelaffen vor (LEVY u. MIRKOVIC 1971; O'BRIEN et al. 1981). Da das Masernvirus, wie andere Paramyxoviridae, hoch kontagiös ist, breitet es sich epizootisch über Aerosole von Tier zu Tier aus. Auch wenn Infektionen vor allem bei Tieren in Gefangenschaft auftreten, sind Serokonversionen für Masernviren ebenfalls in wild lebenden Populationen beschrieben worden. In beiden Fällen scheint jedoch der Mensch im Rahmen einer Zooanthroponose für die initiale Infektion verantwortlich zu sein (BAILEY u. MANSFIELD 2010). Auch hier ist eine Erregerübertragung durch Pflegepersonal wahrscheinlich, da Zukäufe externer Weißbüschelaffen äußert selten sind, und diese Tiere zuerst eine sechswöchige Quarantänezeit durchlaufen, bevor sie in die DPZ eigene Kolonie aufgenommen werden. Die klinischen Symptome und die Pathomorphologie variieren zwischen den unterschiedlichen Spezies sehr stark. Javaner- und auch Rhesusaffen weisen die größten Gemeinsamkeiten mit dem Menschen auf und dienen deshalb als etabliertes 75 Diskussion Tiermodell für Maserninfektionen (EL MUBARAK et al. 2007). Typische Anzeichen, wie Konjunktivitis, makulopapulöse Exantheme und weiße Erosionen der Mundschleimhaut (Koplik-Flecken), sind bei Neuweltaffen kaum beschrieben. Oft kommt es, wie in diesem Fall bei zwei der Tiere, zu subklinischen Verläufen. Nur bei einem Weißbüschelaffen wurde eine progressive Verschlechterung des Allgemeinbefindens protokolliert. Auch die histologischen Befunde unterscheiden sich von denen der Altweltaffen und sind oft weniger stark ausgeprägt, wenngleich Masernviren besonders bei Krallenaffen zu schweren Erkrankungen mit Mortalitätsraten bis zu 100% führen können (LUDLAGE u. MANSFIELD 2003). So wurden schon im Jahr 1966 Krankheitsausbrüche in Krallenaffenkolonien beschrieben, bei denen mit 326 Tieren die Mehrzahl verstarb (LEVY u. MIRKOVIC 1971). Typischerweise neigen Weißbüschelaffen zu nekrotisierenden Entzündungen des Gastrointestinaltraktes, wobei gelegentlich blass eosinophile, zytoplasmatische teils intranukleäre Einschlusskörperchen und Riesenzellen auftreten. Der Magen der positiv getesteten Tiere blieb ohne besonderen Befund. Bei einem Tier zeigte sich, wie in der Literatur beschrieben, in Dünn- und Dickdarm eine chronisch-aktive, teils transmurale lymphozytäre Enteritis mit einer deutlichen Hyperplasie der regionalen Lymphknoten. Bei einem anderen Tier lag eine segmentale mittel- bis hochgradige chronisch-aktive erosive Duodenitis mit deutlicher Zottenatrophie, Krypthyperplasie, multifokalen Kryptabszessen und Schleimhauterosionen vor. Weiterhin zeigte sich ein mittelgradiges propriales gemischtzelliges Entzündungszellinfiltrat mit Beteiligung von neutrophilen Granulozyten und mittelgradiger GALT Hyperplasie. Einschlusskörperchen oder Riesenzellen ließen sich jedoch im Magen-Darm-Bereich nicht feststellen. Neben einer gastrointestinalen Manifestation dieser Erkrankung weist das Masernvirus bei Neuweltaffen ebenfalls einen Gewebetropismus für die Lunge auf. Interstitielle Pneumonien mit Riesenzellen, mononukleären Entzündungszellen, verdickten Alveolarwänden, nekrotische Bereiche mit WarthinFinkelday Zellen und gelegentliche Einschlusskörperchen in den Riesenzellen sowie im Alveolar- oder Bronchialepithel sind charakteristischerweise bei Krallenaffen anzutreffen (LEVY u. MIRKOVIC 1971; LUDLAGE u. MANSFIELD 2003). Auch wenn die molekularbiologischen Ergebnisse des untersuchten Lungengewebes eindeutig 76 Diskussion waren, zeigten sich in unseren Fällen außer einer Alveolarhistiozytose und einer geringgradigen, gemischtzelligen Leukozytostase in den Alveolarsepten keine charakteristischen pathomorphologischen Lungenveränderungen. Wenngleich es sich bei dem Masernnachweis um einen interessanten Befund handelt, kann dieser wegen der geringen Zahl der positiv getesteten Tiere nicht die hohe Prävalenz interstitieller Pneumonien der gesamten Tierkolonie erklären. Ein anderer Erklärungsansatz ergibt sich aus der Untersuchung der Haltungsbedingungen der betroffenen Tiere. Hierbei wurden die Tiere nach dem Ort der Unterbringung in zwei Gruppen unterteilt. Die erste Gruppe bildeten die Weißbüschelaffen, die zwischen Januar 1997 und April 2004 seziert wurden. Alle nach April 2004 geborenen Tiere wurden im neuen Tierhaus gehalten und bildeten die zweite Untersuchungsgruppe. Auffällig ist, dass die Zahl der Weißbüschelaffen mit interstitiellen Pneumonien im Untersuchungszeitraum von 29 % in der alten Haltung um 14 Prozentpunkte auf 43 % in der neuen Haltung angestiegen ist. Diese Unterschiede ließen sich auch statistisch durch einen Chi-Quadrat-Test (p = 0,007) bestätigen, der signifikante Unterschiede zwischen dem Auftreten interstitieller Pneumonien in der neuen und alten Tierhaltung ergab. Unklar ist hingegen, welche Umwelteinflüsse genau das Auftreten interstitieller Pneumonien in der neuen Haltung begünstigten, da sich Hygiene und Haltungsbedingungen zwischen den einzelnen Gebäuden kaum unterschieden. Die Reinigung der Einheiten erfolgte in beiden Tierhäusern einmal wöchentlich durch Hochdruckreiniger. Ebenfalls wurde der Papierbodenbelag unter den Käfigen wöchentlich gewechselt. Es ist bekannt, dass unter bestimmten Klimabedingungen, wie z. B. bei hoher Luftfeuchtigkeit, viele Viren und Bakterien ihre Virulenz länger erhalten können (KEIPER 2002). Daher besteht die Möglichkeit, dass die Weißbüschelaffen besonders nach der Hochdruckreinigung durch die Inhalation des Aerosols einer erhöhten Erregerlast ausgesetzt waren, die Pneumonien begünstigt haben könnte. Die klimatischen Bedingungen variierten zum Untersuchungszeitpunkt nur geringgradig, lagen aber innerhalb der Richtwerte für Temperatur (25 - 26 °C) und Luftfeuchtigkeit (55 - 60 %). Bei stark abfallenden Außentemperaturen können jedoch Temperaturschwankungen besonders in Gebäudeteilen mit Außenwänden auftreten. Einen Unterschied zwischen den beiden 77 Diskussion Tierhäusern stellt die Raumgröße dar. Im alten Gebäude wurden die Weißbüschelaffen in kleineren Räumen von ca.140 m 2 gehalten, wohingegen die Räume im neuen Tierhaus mit 183 m2 größer sind und mit 1000 m3 ein doppelt so großes Volumen aufweisen. Die Luftwechselrate sank nach dem Umzug in die neuen Gebäude von einem 8- auf einen 4,5-fachen Luftwechsel pro Stunde. Chemisch induzierte Pneumonien zählen zu den nichtinfektiösen Ursachen entzündlicher Lungenveränderungen und sind unter anderem auf erhöhte Konzentrationen von Schwefeldioxid (SO2), Stickstoffdioxid, (NO2), Ozon (O3), Hydrogensulfid (H2S) und Feinstaubpartikel in der Umgebungsluft zurückzuführen. Als Folge steigender Luftverschmutzung treten vor allem in urbanen Ballungszentren und Industrieregionen chronische Atemwegserkrankungen wie COPD und Asthma auf (KATSOUYANNI 1995; LE et al. 2012; SANCINI et al. 2012). Aber auch interstitielle Lungenentzündungen sind in diesem Zusammenhang beschrieben (JEND 1995). Die Ursachen der Emissionen sind häufig Verbrennungsprozesse, bei denen eine Vielzahl von Gasen und Partikeln freigesetzt wird, die sich vor allem auf das kardiopulmonale Organsystem auswirken und akute sowie chronische Organveränderungen hervorrufen (BRUNEKREEF et al. 2009). Erhebungen über den Anteil der oben beschriebenen Gase in der Außenluft in Göttingen liegen nicht vor. Nach Informationen der Weltgesundheitsorganisation (WHO) lagen die Feinstaubwerte (PM10) in Göttingen 2011 mit 21 µg pro Kubikmeter leicht über dem empfohlenen Grenzwert von 20 µg. Die als Feinstaub (PM10) bezeichnete Staubfraktion enthält Teilchen, die einen Größen selektierenden Lufteinlass passieren, der für einen aerodynamischen Durchmesser von 10 µm eine Abscheidewirksamkeit von 50 % besitzt. Besonders die kleineren Partikel gelangen bis tief in die Alveolen und sind daher für die gesundheitsschädliche Wirkung verantwortlich. Da die Luftzufuhr zu den Tiereinheiten des DPZ aber durch Filter mit einer Partikelabscheidewirksamkeit bis zu einem Mikrometer reguliert wird, waren die Krallenaffen nur einer geringen Feinstaubbelastung ausgesetzt. Neben den oben genannten Ursachen für Gasemissionen tritt Hydrogensulfid ebenfalls bei Fäulnisprozessen aus Fäkalien aus und führt besonders in der Massentierhaltung, wo die Tiere unter ungünstigen klimatischen Bedingungen direkt mit ihren Fäkalien in 78 Diskussion Berührung kommen, zu respiratorischen Symptomen. Inwieweit auf Einstreu gehaltene Weißbüschelaffen in der Tierhaltung des DPZ einer Belastung mit Hydrogensulfid ausgesetzt sind, ist unklar, dürfte sich vermutlich jedoch in Grenzen halten, da die Tiere als obligate Baumbewohner Bodenkontakt meiden. Unter Berücksichtigung der reduzierten Luftwechselrate in den neuen Tiereinheiten kann eine Anreicherung der Umgebungsluft mit bestimmten sich vor Ort bildenden zytotoxischen Gasen dennoch nicht ausgeschlossen werden und stellt eine mögliche ätiologische Komponente dar. Neben der Art der Unterbringung scheint auch das Alter der Tiere einen Einfluss auf das Auftreten von interstitiellen Lungenentzündungen zu haben. Bei dem Vergleich der drei Altersklassen zeigten sich 51 interstitielle Pneumonien bei neugeborenen, 60 bei juvenilen und 94 bei adulten Weißbüschelaffen. Im Verhältnis zur gesamten Altersgruppe waren mit 42 % besonders juvenile Tiere betroffen, während Neugeborene und Adulte nur zu 30 % bzw. 29 % Veränderungen aufwiesen. Ein signifikanter Unterschied konnte ebenfalls statistisch durch den Chi-Quadrat-Test (p = 0,1256) belegt werden, der eine höhere Wahrscheinlichkeit für das Auftreten interstitieller Pneumonien in der Gruppe der juvenilen Tiere feststellte. Warum gerade diese Tiere eine erhöhte Prävalenz aufweisen, bleibt eine offene Frage. Falls es sich um eine infektiöse Genese handeln sollte, wäre zu vermuten, dass eher Neugeborene nach immunsupprimierte Absinken adulte der Tiere maternalen betroffen Antikörper wären. und Weiterhin andererseits muss davon ausgegangen werden, dass die Veränderungen reversibel sind. Neben den interstitiellen Pneumonien traten am zweithäufigsten bei insgesamt 10 % der untersuchten Tiere Lungenveränderungen auf, die wegen ihrer minimalen Ausprägung nicht als entzündliche Form, sondern rein deskriptiv als minimale mononukleäre interstitielle Zellinfiltrate beschrieben wurden. Die Abgrenzung zu interstitiellen Pneumonien erfolgte über die Septendicke. Immunhistochemische Untersuchungen ergaben, dass neben Makrophagen vor allem T-Lymphozyten zu einem Anstieg der Septendicke bis 30 µm führten. Die Mehrzahl der Tiere wies einen subakuten Krankheitsverlauf und eine vorwiegend multifokale Verteilung auf. Statistische Untersuchungen stellten keine Signifikanzen für das Geschlecht oder 79 Diskussion das Alter fest. Über das Auftreten von minimalen Entzündungszellinfiltraten im Lungeninterstitium bei Weißbüschelaffen liegen in der Literatur keine Berichte vor. Klinisch treten diese Veränderungen, wie auch die interstitiellen Pneumonien, nicht in Erscheinung, sodass zu vermuten ist, dass sie die Tiere nicht beeinträchtigen. Möglicherweise handelt es sich bei diesen minimalen Infiltraten um eine Vorstufe der interstitiellen Pneumonien oder um ein unspezifisches Reaktionsmuster auf bestimmte Haltungsbedingungen und sind als gelegentlich auftretende „background findings“ zu interpretieren. Bei 18 Weißbüschelaffen (3 % der Tiere mit Lungenveränderungen) wurden histologisch alveoläre Pneumonien diagnostiziert. Anders als bei den interstitiellen Pneumonien und minimalen septalen Zellinfiltraten zeigten die Tiere hier je nach Schweregrad vereinzelt klinische Symptome. Anamnestisch wurden Apathie und Beeinträchtigungen im Respirationstrakt bei drei Weißbüschelaffen vermerkt. Während die interstitiellen Pneumonien in keinem der Fälle die alleinige Todesursache darstellten, starben 14 der 18 Tiere an der alveolären Pneumonie. Streptococcus sp. und Bordetella bronchiseptica konnten mikrobiologisch aus dem Lungengewebe der 6 Tiere mit Bronchopneumonien isoliert werden. Nach einem gehäuften Auftreten von eitrigen Lungenentzündungen im Bestand der Krallenaffen im Sommer 1998 wurden und werden die Tiere ab einem Alter von drei Monaten routinemäßig mit stallspezifischen Vakzinen gegen Bordetellen und mit handelsüblichen Impfstoffen gegen Rotlauf und Yersiniose immunisiert. Bei den hier erwähnten Weißbüschelaffen mit Bordetellose handelt es sich um juvenile Tiere, die, noch bevor sie geimpft wurden, an den Folgen der Pneumonie verstarben. Auch bei den neun Lobärpneumonien, die in sechs Fällen die Todesursache darstellten, wies ein zwei Monate altes Tier ebenfalls eine Infektion mit Bordetella bronchiseptica auf. Des Weiteren wurden Streptokokken, Enterokokken und Klebsiella pneumoniae sp. ozaenae nachgewiesen. Die weniger pathogenen Enterokokken zählen zur normalen Darmflora, treten als opportunistische Infektionserreger in Erscheinung und sind in der Humanmedizin an nosokomialen Infektionen beteiligt. Streptokokken, Klebsiellen und Bordetellen wurden im Respirationstrakt gesunder nicht humaner Primaten nachgewiesen, wobei Krankheitsausbrüche häufig in Verbindung mit Tiertransporten, 80 Diskussion Quarantäne oder Überbelegung standen (ABEE et al. 2012; SOLTANI et al. 2012). In einem weiteren Fall gelang zwar keine kulturelle Anzucht, aufgrund der histologischen Ausprägung wurde aber vermutet, dass es sich bei den floriden Herden mit dichten Bakterienkolonien um eine Aktinomykose oder Nokardiose handelte. Bei einem Tier konnte histologisch unter der Verwendung von Spezialfärbungen eine Lungencandidose diagnostiziert werden. Für den Erregereintrag in die Kolonie kommen verschiedene Möglichkeiten infrage. Sehr wahrscheinlich dürften die Tiere mit infiziertem Futter (z. B. Obst oder Insekten) in Kontakt gekommen sein, aber auch eine aerogene Infektion durch die Außenluft ist in Betracht zu ziehen. Zwar durchströmt die angesaugte Luft einen Feinstaubfilter, aber dieser hält effektiv nur Partikel ab einer Größe von 3 µm zurück. Daher können durch die Luft Viren und die Mehrzahl der Bakterien ungehindert in das Tierhaus gelangen. Ein Zukauf von externen Tieren blieb im Untersuchungszeitraum aus, sodass dieser Übertragungsweg ausgeschlossen werden kann. Nicht ausgeschlossen, aber weniger wahrscheinlich, ist die Übertragung durch Tierpfleger oder Tierärzte. Zugangsbeschränkungen, Schleusenbereiche mit Duschen und Kleiderwechsel und engmaschige betriebsärztliche Untersuchungen sollten den Erregereintrag durch den Menschen so gering wie möglich halten. Fünf Tiere wiesen neoplastische Veränderungen der Lunge auf. Dazu gehörten vier Lymphome und ein Fibrosarkom. Für Krallenaffen liegen zahlreiche Fallberichte über Lymphomerkrankungen vor (HOFFMANN et al. 2001) und in Untersuchungen von DAVID et al. (2009) mit 129 Weißbüschelaffen wird das maligne Lymphom als die am häufigsten auftretende Tumorerkrankung beschrieben. Ätiologisch kommt unter anderem eine Infektion mit dem zur Gruppe der Gammaherpesvirinae gehörende Lymphocryptovirus in Betracht. Dieses wurde im Jahr 2000 von RAMER et al. (2000) aus B-Zell-Lymphomen des Weißbüschelaffen isoliert und 2002 vollständig sequenziert (RIVAILLER et al. 2002). Das Virus wurde seitdem bei erkrankten und gesunden Tieren nachgewiesen und serologische Prävalenzen von 75 – 81 % lassen eine starke Durchseuchung der Bestände vermuten (JENSON et al. 2002). Da jedoch trotz der deutlichen Anzahl seropositiver Tiere nur wenige an Lymphomen erkranken, ist anzunehmen, dass die Tumorinduktion zusätzlich durch Faktoren wie 81 Diskussion Stress oder Immunsuppression gefördert wird ähnlich wie bei der Knudsonhypothese (RAMER et al. 2000). Bei dieser ging Alfred Knudson 1971 davon aus, dass die Ausbildung von Tumoren im Rahmen einer „two-hit“ Hypothese dann besonders früh stattfindet, wenn eines der beiden Tumorsuppressorgene durch einen genetischen Defekt bereits deaktiviert ist (MCGAVIN u. ZACHARY 2007). Zu den häufigsten Befunden der „side effects“, also subklinischen sekundären Lungenveränderungen, die nicht in direktem Kontakt zum Krankheitsgeschehen stehen, zählten Verkalkungen, die zufällig über das Lungenparenchym verteilt waren. Mineralisationen sind beim Weißbüschelaffen in verschiedenen Organen beschrieben (KASPAREIT et al. 2007). Berichte über das Auftreten in der Lunge existieren bei dieser Tierart nicht. Von Lungenparenchymnekrosen ausgehende dystrophische Verkalkungen hyperkalzämischen Zuständen oder sind metastatische mögliche Mineralisationen Ursachen. Auch wenn bei bei stichprobenartigen Untersuchungen der Tiere mit Lungenverkalkungen keine weiteren Mineralisationen in den übrigen Organen festgestellt werden konnten, sind dystrophische Ursachen wenig wahrscheinlich, da keine Anzeichen für degenerative oder nekrotische Prozesse vorlagen. Für metastatische Mineralisationen spricht die Tatsache, dass dem Tierfutter Vitamin D3 zugesetzt wird, um einem Mangel an UVStrahlung in der Indoor-Haltung auszugleichen. Bei Weißbüschelaffen ist bekannt, dass sie, verglichen mit dem Menschen und anderen nicht humanen Primaten, eine bis zu zehnfach höhere Cholecalciferol-Konzentration im Blut aufweisen (SHINKI et al. 1983). Um diesen Ansprüchen gerecht zu werden und um Knochendeformationen und Wachstumsstörungen vorzubeugen, werden daher täglich pro Tier 0,008 mg Ursovit® (Fa. Rebopharm, Bocholt) und 0,18 g Vitamin D3 reiches, speziell angefertigtes Mineralstoffpulver (Fa. Kawo, Hildesheim) supplementiert. Eine leichte Überdosierung führte vermutlich zu einem deutlich erhöhten Kalziumspiegel im Blut, sodass sich dieses an kollagen- und elastinreichem Gewebe, wie z. B. alveolären Septen, Gefäßwänden oder Bronchiolenwänden, ablagerte und die beschriebenen runden bis ovalen Mineralisationsherde bildete. Die pulmonale Anthrakose ist ein häufiger Zufallsbefund bei Tieren, die in industrialisierten Gebieten gehalten werden (CASWELL u. WILLIAMS 2008). Da die 82 Diskussion Weißbüschelaffen in reinen Indoor-Haltungen untergebracht sind und die Luft einen Filter durchströmt, der alle Rußpartikel über 3 µm abscheidet, ist die Belastung mit Kohlenstoffpigment in der Tierhaltung des DPZ als weniger stark zu betrachten. Fallberichte für Anthrakose liegen vor allem für Makaken vor, die Zugang zu Außenbereichen haben (YAMATE et al. 2009). Untersuchungen von SHIMOI et al. (1998) wiesen Anthrakose bei allen der 166 in Labortierhaltung untergebrachten Javaneraffen nach, wohingegen nur 10 % der 161 Tiere der wild lebenden Vergleichspopulation betroffen waren. Aktuelle humanmedizinische Veröffentlichungen berichten von einem gehäuften Auftreten der Anthrakose bei Tuberkulosepatienten (PAZOKI et al. 2012). In der Tiermedizin liegen dazu keine Erkenntnisse vor. Der Nachweis von Hämosiderin war im Untersuchungszeitraum rückläufig. Neben kardial bedingten Stauungen und einer daraus resultierenden Diapedese der Erythrozyten in die Alveolen ist ebenfalls die alimentäre Aufnahme von Eisen ein bedeutender Faktor für die Bildung und pulmonale Ablagerung dieses Pigments. Neben Einzelfällen, bei denen Herzfehlerzellen nachgewiesen werden konnten, ist zu vermuten, dass in der Vergangenheit die Mehrzahl der Hämosiderosen auf einer alimentären Eisenüberversorgung resultierte. Mit der Umstellung des Futters 2006 auf eine eisenärmere Diät traten Hämosiderinablagerungen seltener auf. In dieser Dissertation wurde retrospektiv das Vorkommen pathologischer Veränderungen im Respirationstrakt des Weißbüschelaffen untersucht. Weiterhin sollten anhand aktueller Sektionsfälle mithilfe von Kulturverfahren und molekularbiologischen Methoden die Beteiligung möglicher infektiöser Erreger abgeklärt werden. Betrachtet man die Ergebnisse dieser Untersuchung, so erscheinen folgende Punkte wesentlich für Weißbüschelaffen, die in Tierversuchen eingesetzt werden sollen. Genaue Bestimmung des Gesundheitszustandes vor Versuchsbeginn Im Rahmen einer Allgemeinuntersuchung sollte vor Versuchsbeginn der genaue Gesundheitszustand des Tieres festgestellt werden. Dazu zählt das 83 Diskussion Erfassen der Körperinnentemperatur, die Auskultation und die Bestimmung des Blutbildes, um Hinweise für Entzündungsprozesse (Kernlinksverschiebung/ Leukozytostase) zu erhalten. Anzeichen alveolärer Pneumonien sind feinblasige Rasselgeräusche bei der Auskultation, Husten mit eitrigem Auswurf oder eine erhöhte Anzahl neutrophiler Granulozyten in der Spülflüssigkeit der bronchoalveolären Lavage. In dieser Untersuchung wurden solche Symptome nicht beschrieben, da hier die Tiere mit Lungenveränderungen fast ausschließlich interstitielle Pneumonien oder minimales mononukleäres Infiltrat in den Septen zeigten. Sofern diese Lungenveränderungen nicht erst bei der Sektion, sondern am lebenden Tier festgestellt werden sollen, gestaltet sich die Diagnostik als äußert schwierig. Interstitielle Lungenentzündungen lassen sich, abhängig vom Schweregrad und der Ausprägung, mit bildgebenden Verfahren, wie z. B. Röntgenuntersuchungen, darstellen. Um sicher disseminierte Pneumonien ausschließen zu können, empfiehlt sich die Biopsie mittels Bronchoskopie, um das Gewebe histologisch zu untersuchen. Interpretation der histologischen Befunde im Rahmen von Tierversuchen Selbst mit einer aufwendigen Diagnostik stellt die präzise Feststellung des Gesundheitszustandes der Lunge, vor allem bei minimalen Infiltraten in den Septen oder bei interstitiellen Pneumonien, eine besondere Herausforderung dar. Daher ist bei der Erhebung histologischer Befunde am Versuchsende zu bedenken, dass mitunter auftretende Veränderungen bereits vorher schon bestanden und nicht diagnostiziert werden konnten. Diese pathomorphologischen Befunde, die nicht in direktem Zusammenhang mit dem Tierversuch oder Erkrankungen stehen und als unspezifische Begleiterscheinungen auftreten, sind im Rahmen von „background findings“ zu interpretieren. 84 Diskussion Für die Weißbüschelaffenkolonie des DPZ ergab diese Untersuchung, dass es sich bei den Befunden in der Lunge vor allem um interstitielle Pneumonien und minimale mononukleäre, septale Infiltrate handelte. Da weder histologisch noch molekularoder mikrobiologische Analysen einen Hinweis auf infektiöse Ätiologie ergaben, muss von nichtinfektiösen Ursachen ausgegangen werden. Besondere Beachtung sollte vor Ort entstandenen Umweltbelastungen geschenkt werden, die sich durch reduzierte Luftwechselraten in den neuen Tiereinheiten anreichern könnten. Daher ist zu empfehlen, die klimatischen Bedingungen in den neuen Tiereinheiten eingehend zu untersuchen und gegebenenfalls anzupassen. Auch über die Möglichkeit einer Außenhaltung ist nachzudenken, obwohl bei dieser andere Pathogenbelastungen unter Umständen negative Einflüsse haben. 85 Zusammenfassung 6 Zusammenfassung Marius Kunze (2012) Retrospektive Untersuchungen zu Lungenveränderungen bei Weißbüschelaffen (Callithrix jacchus) in einer Versuchstierkolonie. Im Rahmen einer Kooperation zwischen dem Fraunhofer Institut für Toxikologie und Experimentelle Medizin (ITEM), Hannover und dem Deutschen Primatenzentrum (DPZ), Göttingen existiert ein Projekt, in dem der Weißbüschelaffe als Tiermodell für chronische Atemwegserkrankungen des Menschen, u. a. COPD und Asthma, etabliert werden soll. Das Ziel dieser Arbeit war, das Spektrum der Lungenpathologie beim Weißbüschelaffen zu definieren, um pathologische Prozesse von möglichen „background lesions“ abzugrenzen. Ein weiterer Untersuchungsschwerpunkt lag in der Bestimmung gesundheitsrelevanter Pathogene des Respirationstraktes beim Weißbüschelaffen, deren Kenntnis der Gewährleistung eines definierten Gesundheitsstatus der für Versuche verwendeten Tiere dient. Retrospektiv wurde im Zeitraum von 1997 bis 2011 bei insgesamt 639 Tieren im Bestand der Weißbüschelaffenkolonie des DPZ das Spektrum von Lungenveränderungen mit Schwerpunkt auf entzündlichen Lungenveränderungen untersucht. Um die gewonnenen Erkenntnisse besser vergleichen zu können, wurden zwei weitere externe Tiergruppen in diese Untersuchung aufgenommen. Dabei handelte es sich um 38 Weißbüschelaffen der Universität Göttingen und um 34 Tiere einer externen Labortiereinrichtung. Die pathomorphologischen Diagnosen wurden zusammen mit den Ergebnissen der ätiologischen Untersuchungen (Mikrobiologie, Parasitologie) und tierspezifischen Informationen (Alter, Geschlecht, Vorbericht, Haltung) in Excel-Tabellen überführt und statistisch ausgewertet. Die Analysen ergaben, dass die Prävalenz für Pneumonien im Bestand des DPZ bei 35 % lag. Der Vergleich mit den ebenfalls ausschließlich indoor gehaltenen Tieren der externen Labortiereinrichtung ergab mit 53 % einen deutlich höheren Wert. Die Universitätstiere mit Zugang zu einem Außenbereich zeigten lediglich eine Prävalenz von 5 %. Die vorliegende Studie ergab, dass es sich bei den hauseigenen Tieren mit 86 Zusammenfassung Lungenveränderungen vor allem um interstitielle Pneumonien (32 % der untersuchten Tiere) und um minimale Entzündungszellinfiltrate (10 % der untersuchten Tiere) handelte. Hinweise auf Infektionserreger fanden sich weder histologisch noch kulturell bakteriologisch. Statistische Untersuchungen zum Auftreten von minimalen Entzündungszellinfiltraten ergaben keine Geschlechts- oder Altersprädisposition. Signifikanzen zeigten sich jedoch bei den interstitiellen Pneumonien. Zum einen waren vor allem juvenile Tiere (10 Wochen bis 30 Monate) betroffen und zum anderen traten diese unterschiedlich oft in den beiden Tierhäusern des DPZ auf. Ein geringer Anteil der untersuchten DPZ-Tiere (4 %) wies alveoläre Pneumonien, bronchointerstitielle Pneumonien, granulomatöse Entzündungen oder Lungentumore auf. Ätiologisch konnten verschiedene Infektionserreger wie Streptokokken oder Bordetellen nachgewiesen werden. Darüber hinaus wurde anhand aktueller Sektionsfälle bei 22 DPZ-Tieren mithilfe molekularbiologischer Methoden versucht, Hinweise auf eine mögliche infektiöse Ätiologie der entsprechenden interstitiellen Läsionen zu erhalten, insbesondere im Hinblick auf eine Beteiligung viraler Pathogene, wie das Respiratorische Synzytialvirus A und B, Influenza A, Influenza B, Adenoviren, Enteroviren, Picornaviren, Pan Paramyxoviren und Masern. Bei zwei Tieren gelang der Nachweis des Maserngenoms aus tiefgefrorenem Lungengewebe. Die geringe Zahl der positiv getesteten Tiere kann nicht die hohe Prävalenz interstitieller Pneumonien der gesamten Tierkolonie erklären. Wahrscheinlicher ist es, dass deren Auftreten mit haltungsspezifischen Faktoren in Zusammenhang steht, zumal sich diese in Bezug auf die Luftwechselrate zwischen beiden Tierhäusern unterscheiden. In den neuen Tiereinheiten sollten daher die klimatischen Bedingungen untersucht und gegebenenfalls angepasst werden. Für in Versuchen für respiratorische Erkrankungen verwendete Weißbüschelaffen ergeben sich aus dieser Untersuchung folgende Aspekte. Vor Versuchsbeginn ist eine genaue Bestimmung des Gesundheitszustandes essenziell, um eventuelle vorliegende Pneumonien zu diagnostizieren, die auf einer infektiösen Genese beruhen. Der Ausschluss von interstitiellen Pneumonien oder 87 Zusammenfassung minimalen Entzündungszellinfiltraten in den Septen kann allerdings, insbesondere bei einem nicht infektiösen Ursprung, eine besondere Herausforderung darstellen und wird oftmals nur histologisch möglich sein. Bei der Interpretation histologischer Befunde im Rahmen von Tierversuchen ist das Auftreten interstitieller entzündlicher Lungenveränderungen zu beachten und im Sinne von „background lesions“ von versuchsbedingten Veränderungen zu unterscheiden. 88 Summary 7 Summary Marius Kunze (2012) Retrospective investigation of lung changes in marmosets (Callithrix jacchus) in a laboratory animal facility. The common marmoset is employed as an animal model for chronic respiratory diseases in humans – specifically chronic obstructive pulmonary disease (COPD) and asthma – in a collaborative research effort between the Fraunhofer Institute for Toxicology and Experimental Medicine (ITEM), Hannover and the German Primate Center (DPZ), Göttingen. The study presented in this thesis defines the spectrum of lung pathology in common marmosets with the express purpose of delimiting pathological processes of possible "background lesions" in experimental animals. Moreover, a fundamental element of the investigation lies in determining the healthrelated respiratory pathogens in marmosets. Another main focus of the study was the determination of health-relevant pathogens in the respiratory system of marmosets. The results obtained from this approach can be used in future research studies. This study, between 1997 and 2011 in a total of 639 animals in the care of the marmoset colony of DPZ, investigated the spectrum of lung changes with an emphasis on inflammatory lung lesions. To better compare the findings, two additional external groups of animals were included in this study: 38 marmosets from the University of Göttingen and 34 animals from an external laboratory animal facility. Pathomorphological diagnoses were combined with the results of etiological studies (microbiology, parasitology) and animal-specific information (age, gender, preliminary report, housing conditions) into Excel spreadsheets and analyzed statistically. The analysis showed that the prevalence of pneumonia in the DPZ group (housed exclusively indoors) was 35%. In comparison, the prevalence of pneumonia in animals of the external facility, which were also housed exclusively indoors, was much higher at 53%. The animals held at the university, with access to an outdoor area, showed a prevalence rate of 5%. The results from the present study show that, 89 Summary 32% of the tested in-house animals had interstitial pneumonia while only 10% presented with inflammatory cell infiltrate. With regard to infectious agents, neither histological nor cultural bacteria were found. Statistical surveys for the presence of sparse inflammatory cell infiltrate showed no gender or age predisposition. Significant findings, however, lay in the cases of interstitial pneumonia. Firstly, juvenile animals (age 10 weeks to 30 months) were most affected. Furthermore, the occurrence of interstitial pneumonia was different between the two animal houses of the DPZ (i.e. new facility vs. old facility). A small proportion of the animals (4%) studied in the DPZ group exhibited alveolar pneumonia, bronchointerstitial pneumonia, granulomatous inflammation or lung tumors. Etiologically, various pathogens such as streptococcus or bordetella could be detected. This study also aimed to obtain information on the possible infectious etiology of the corresponding interstitial lesions by utilizing current sections cases in 22 DPZ animals and molecular biological methods. Specific emphasis was placed on the possible participation of viral pathogens, such as respiratory syncytial virus A and B, influenza A, influenza B, adenoviruses, enteroviruses, picornaviruses, panparamyxoviruses and measles. The measles genome from frozen lung tissue was detected in two animals. This small number of positive tested animals cannot account for the high prevalence of interstitial pneumonia in the entire animal colony. One likely hypothesis is that it occurs with housing-specific factors, since they are different in terms of the air exchange rate within the two animal houses. Therefore, the climatic conditions should be examined and adjusted in the new animal units, if necessary. The results from the present study emphasize an important consideration prior to conducting studies on respiratory diseases in marmosets. Before experiments, an accurate and thorough determination of the health status of an animal is essential in order to diagnose any present pneumonia based on an infectious etiology. The exclusion of interstitial pneumonia or minimal inflammatory cell infiltration in the septa 90 Summary may, however, represent a particular challenge, especially in cases of non-infectious origin; this diagnosis will often only be histologically possible. When interpreting histologic findings in the context of animal experiments, it is important to consider the occurrence of preexisting inflammatory interstitial disease and to distinguish previous background lesions from trial-related changes. 91 Literaturverzeichnis 8 Literaturverzeichnis ABBOTT, D. H., D. K. BARNETT, R. J. COLMAN, M. E. YAMAMOTO u. N. J. SCHULTZ-DARKEN (2003): Aspects of common marmoset basic biology and life history important for biomedical research. Comp Med 53, 339-350 ABEE, C. R., K. MANSFIELD, S. D. TARDIF u. A. MORRIS (2012): Nonhuman Primates in Biomedical Research: Diseases. 2. Auflage, Elsevier ALBRECHT, P., D. LORENZ, M. J. KLUTCH, J. H. VICKERS u. F. A. ENNIS (1980): Fatal measles infection in marmosets pathogenesis and prophylaxis. Infect Immun 27, 969-978 ANDRADE, M. C. u. R. S. MARCHEVSKY (2007): Histopathologic findings of pulmonary acariasis in a rhesus monkeys breeding unit. Rev Bras Parasitol Vet 16, 229-234 BAILEY, C. u. K. MANSFIELD (2010): Emerging and reemerging infectious diseases of nonhuman primates in the laboratory setting. Vet Pathol 47, 462-481 BASKERVILLE, M., M. WOOD u. A. BASKERVILLE (1983): An outbreak of Bordetella bronchiseptica pneumonia in a colony of common marmosets (Callithrix jacchus). Lab Anim 17, 350-355 BELSHE, R. B., L. S. RICHARDSON, W. T. LONDON, D. L. SLY, J. H. LORFELD, E. CAMARGO, D. A. PREVAR u. R. M. CHANOCK (1977): Experimental respiratory syncytial virus infection of four species of primates. J Med Virol 1, 157-162 BENJAMIN, S. A. u. C. M. LANG (1971): Acute pasteurellosis in owl monkeys (Aotus trivirgatus). Lab Anim Sci 21, 258-262 BERENDT, R. F., G. L. KNUTSEN u. M. C. POWANDA (1978): Nonhuman primate model for the study of respiratory Klebsiella pneumoniae infection. Infect Immun 22, 275-281 BERENDT, R. F., G. G. LONG u. J. S. WALKER (1975): Influenza alone and in sequence with pneumonia due to Streptococcus pneumoniae in the squirrel monkey. The Journal of infectious diseases 132, 689-693 92 Literaturverzeichnis BLEIER, T., U. HETZEL , C. BAUER, O. BEHLERT, E. BURKHARDT (2005): Gongylonema pulchurum infection and esophageal squamous cell carcinoma in a vari (Lemur macaco variegata). J Zoo Wildl Med 36, 342-345 BÖCKER, W. (2008): Repetitorium Pathologie Elsevier, München , Jena BORJI, H., M. EMAMI, M. MALEKI, G. RAZMI, H. KAZEMI MEHRJERDI u. E. MOGHADDAS (2012): Alveolar Echinococcosis Infection in a Monkey (Ateles geoffroyi) in Mashhad, Iran. Iran J Pub Health 2, 111-116 BOYCE, J. T., W. E. GIDDENS, JR. u. M. VALERIO (1978): Simian adenoviral pneumonia. Am J Path 91, 259-276 BRACK, M., E. GUNTHER, H. GILHAUS, W. SALZERT u. J. MEUTHEN (1997): An outbreak of Streptococcus equi ssp. zooepidemicus infection of probable human origin in Wanderoos (Macaca silenus)-case report. Zentralbl Bakteriol 286, 441-446 BRACK, M., P. SCHWARTZ, T. HEINRICHS, M. SCHULTZ u. E. FUCHS (1996): Tumors of the respiratory tract observed at the German Primate Center, 1978-1994. J Med Primatol 25, 424-434 BRUNEKREEF, B., R. BEELEN, G. HOEK, L. SCHOUTEN, S. BAUSCH-GOLDBOHM, P. FISCHER, B. ARMSTRONG, E. HUGHES, M. JERRETT u. P. VAN DEN BRANDT (2009): Effects of long-term exposure to traffic-related air pollution on respiratory and cardiovascular mortality in the Netherlands: the NLCS-AIR study. Res Rep Health Eff Inst 139, 5-71 Lepenies, J (2004) Entzündung In: BÜHLING, K., J. LEPENIES u. K. WITT (Hrsg.): Allgemeine und spezielle Pathologie Urban & Fischer, München, S. 56 BURROWS, A. M. u. L. T. NASH (2010): The Evolution of Exudativory in Primates. Springer, New York CALCEDO, R., L. H. VANDENBERGHE, S. ROY, S. SOMANATHAN, L. WANG u. J. M. WILSON (2009): Host immune responses to chronic adenovirus infections in human and nonhuman primates. J Virol 83, 2623-2631 93 Literaturverzeichnis CASWELL, J. L. u. K. J. WILLIAMS (2008): Respiratory system. In: K. V. F. Jubb, P. C. Kennedy, N. C. Palmer (Hrsg.): Pathology of domestic animals Saunders Elsevier, Academic Press, London, S. 523-655 CHAKRABORTY, A. u. P. GOSWAMI (1996): Pulmonary mycotic infection in non-human primates in Assam state zoo. Ind J Vet Pathol 20, 133-135 CHALMERS, D. T., L. B. MURGATROYD u. P. F. WADSWORTH (1983) A survey of the pathology of marmosets (Callithrix jacchus) derived from a marmoset breeding unit Lab Animal 17, 270-279 CHEN, E. C., S. YAGI, K. R. KELLY, S. P. MENDOZA, R. P. TARARA, D. R. CANFIELD, N. MANINGER, A. ROSENTHAL, A. SPINNER, K. L. BALES, D. P. SCHNURR, N. W. LERCHE u. C. Y. CHIU (2011): Cross-species transmission of a novel adenovirus associated with a fulminant pneumonia outbreak in a new world monkey colony. PLoS Pathog 7, e1002155 CHEN, Y., W. DENG, H. ZHU, J. LI, Y. XU, X. DAI, C. JIA, Q. KONG, L. HUANG, Y. LIU, C. MA, C. XIAO, Q. LI, E. BEZARD u. C. QIN (2009): The pathologic features of neurocutaneous melanosis in a cynomolgus macaque. Vet Pathol 46, 773-775 CHI, F., M. LEIDER, F. LEENDERTZ, C. BERGMANN, C. BOESCH, S. SCHENK, G. PAULI, H. ELLERBROK u. R. HAKENBECK (2007): New Streptococcus pneumoniae clones in deceased wild chimpanzees. J Bacteriol 189, 6085-6088 CHMIELEWICZ, B., J. BENZLER, G. PAULI, G. KRAUSE, F. BERGMANN u. B. SCHWEIGER (2005): Respiratory disease caused by a species B2 adenovirus in a military camp in Turkey. J Med Virol 77, 232-237 CHOI, Y. K., M. A. SIMON, D. Y. KIM, B. I. YOON, S. W. KWON, K. W. LEE u. I. B. SEO (1999): Fatal measles virus infection in Japanese macaques (Macaca fuscata). Vet Pathol 36, 594-600 CHOUDARY, C., M. MOHAN RAO u. S. ALI (1986): Anthracosis in zoo animals and birds. Ind Vet J 10, 869-870 COSTA, E. O., L. S. DINIZ, C. F. NETTO, C. ARRUDA u. M. L. DAGLI (1994): Epidemiological study of sporotrichosis and histoplasmosis in captive Latin American wild mammals, Sao Paulo, Brazil. Mycopathologia 125, 19-22 94 Literaturverzeichnis COSTABEL, U. (2000): The new classification of idiopathic interstitial pneumonia. Pneumologie 54, 447-453 DA CRUZ, J., P. A., R. DA ROCHA SILVA u. A. COIMBRA FILHO (1993): Histoplasmosis in Callithrix geoffroyi (Callitrichidae - Primates). Primatol do Brazil 4, 205-213 DACIC, S. u. S. A. YOUSEM (2003): Histologic classification of idiopathic chronic interstitial pneumonias. Am J Respir Cell Mol Biol 29, S5-9 DAHME, E. u. E. WEISS (2007): Grundriss der speziellen pathologischen Anatomie der Haustiere. Enke Verlag, Stuttgart DAVID, J. M., E. J. DICK, JR. u. G. B. HUBBARD (2009): Spontaneous pathology of the common marmoset (Callithrix jacchus) and tamarins (Saguinus oedipus, Saguinus mystax). J med Primatol 38, 347-359 DICK, E. C. u. C. R. DICK (1974): Natural and experimental infections of nonhuman primates with respiratory viruses. Lab Anim Sci 24, 177-181 DUNCAN, M., L. TELL, C. H. GARDINER u. R. J. MONTALI (1995): Lingual gongylonemiasis and pasteurellosis in Goeldi´s monkeys (Callimico goeldii). J Zoo Wildl Med 26, 102-108 DURBIN, A. P., W. R. ELKINS u. B. R. MURPHY (2000): African green monkeys provide a useful nonhuman primate model for the study of human parainfluenza virus types-1, -2, and -3 infection. Vaccine 18, 2462-2469 DZHIKIDZE, Z. K., R. I. KRYLOVA, N. A. VOSKANIAN, G. G. KUKAVA u. P. DZHINDZHIIA (1996): [An outbreak of Streptococcus zooepidemicus-caused infection among laboratory primates]. Zh Mikrobiol Epidemiol Immunobiol 1, 81-84 EDMAN, J. C., J. A. KOVACS, H. MASUR, D. V. SANTI, H. J. ELWOOD u. M. L. SOGIN (1996) An outbreak of Dtreptococcus zooepidemicus-caused infection among laboratory primates. Zh Mikrobiol Epidemiol Imunobiol 1, 81-84 EL MUBARAK, H. S., S. YUKSEL, G. VAN AMERONGEN, P. G. MULDER, M. M. MUKHTAR, A. D. OSTERHAUS u. R. L. DE SWART (2007): Infection of cynomolgus macaques (Macaca fascicularis) and rhesus macaques (Macaca mulatta) with different wild-type measles viruses. J Gen Virol 88, 2028-2034 95 Literaturverzeichnis ETS 123 (European Treaty Serie 123) (2007): Empfehlung der Kommission mit Leitlinien für die Unterbringung und Pflege von Tieren, die für Versuche und andere wissenschaftliche Zwecke verwendet werden. Amtsblatt der Europäischen Union AZ K(2007) 2525 FLECKNELL, P. A., R. PARRY, J. R. NEEDHAM, R. M. RIDLEY, H. F. BAKER u. P. BOWES (1983): Respiratory disease associated with parainfluenza Type I (Sendai) virus in a colony of marmosets (Callithrix jacchus). Lab Anim 17, 111-113 FLYNN, J. L., S. V. CAPUANO, D. CROIX, S. PAWAR, A. MYERS, A. ZINOVIK u. E. KLEIN (2003): Non-human primates: a model for tuberculosis research. Tuberculosis (Edinb) 83, 116-118 FORTMAN, J. D., J. R. MANALIGOD u. B. T. BENNET (1993): Malignant mesothelioma in an olive baboon (Papio anubis). Lab Anim Sci 43, 503-505 FOX, J. G., B. J. COHEN u. L. F. M. (1984): Laboratory Animal Medicine. Academic Press, Inc., Orlando, Folrida GARCIA, A., P. R. NAMBIAR, R. P. MARINI u. J. G. FOX (2009): Staphylococcal meningoencephalitis, nematodiasis, and typhlocolitis in a squirrel monkey (Saimiri sciureus). Journal of medical primatology 38, 377-381 GERN, J. E. (2010): The ABCs of rhinoviruses, wheezing, and asthma. J Virol 84, 7418-7426 GRAYBILL, J. R., L. GRIFFITH u. S. H. SUN (1990): Fluconazole therapy for coccidioidomycosis in Japanese macaques. Rev Infect Dis 12, 286-290 GREENOUGH, T. C., A. CARVILLE, J. CODERRE, M. SOMASUNDARAN, J. L. SULLIVAN, K. LUZURIAGA u. K. MANSFIELD (2005): Pneumonitis and multi-organ system disease in common marmosets (Callithrix jacchus) infected with the severe acute respiratory syndrome-associated coronavirus. Am J Pathol 167, 455-463 GUSTAVSSON, O. E., B. O. ROKEN u. R. SERRANDER (1990): An epizootic of whooping cough among chimpanzees in a zoo. Folia Primatol (Basel) 55, 45-50 96 Literaturverzeichnis HAGEDORN, M. (1977) Lunge In: SANDRITTER, W., C. THOMAS, N. BÖHM, N. FREUDENBERG, M. HAGEDORN, N. RIEDE u. K. SALFELDER (Hrsg.): Histopathologie - Lehrbuch und Atlas für Studierenede und Ärzte. F. K. Schattauer Verlag, Stuttgart; S. 83-111 HAIG, D. (1999): What is a marmoset? Am J Primatol 49, 285-296 HALL, W. C., R. M. KOVATCH, P. H. HERMAN u. J. G. FOX (1971): Pathology of measles in rhesus monkeys. Vet Pathol 8, 307-319 HARRISON, M. L. u. S. D. TARDIF (1994): Social implications of gummivory in marmosets. Am J Phys Anthropol 95, 399-408 HAYDEN, F. G. (2004): Rhinovirus and the lower respiratory tract. Rev Med Virol 14, 17-31 HEARN, J. P. (1983): Reproduction in new world primates: New model in medical science. MTP Press, Boston HERFST, S., J. M. VAN DEN BRAND, E. J. SCHRAUWEN, E. DE WIT, V. J. MUNSTER, G. VAN AMERONGEN, M. LINSTER, F. ZAARAOUI, W. F. VAN IJCKEN, G. F. RIMMELZWAAN, A. D. OSTERHAUS, R. A. FOUCHIER, A. C. ANDEWEG u. T. KUIKEN (2010): Pandemic 2009 H1N1 influenza virus causes diffuse alveolar damage in cynomolgus macaques. Vet Pathol 47, 1040-1047 HERRIN, K. V., A. MIRANDA u. D. LOEBENBERG (2005): Posaconazole therapy for systemic coccidioidomycosis in a chimpanzee (Pan troglodytes): a case report. Mycoses 48, 447-452 HESSLER, J. R. u. A. F. MORELAND (1968): Pulmonary tuberculosis in a squirrel monkey (Saimiri sciureus). J Am Vet Med Assoc 153, 923-927 HILL, D. u. J. P. DUBEY (2002): Toxoplasma gondii: transmission, diagnosis and prevention. Clin Microbiol Infect 8, 634-640 Hofmann, P., K. Kahnt, K. Mätz-Rensing, M. Brack, F. J. Kaup (2001) Three spontaneous lymphomas in a colony of cotton-top tamarins (Saguinus oedipus). J Med Primatol 6, 322-7. 97 Literaturverzeichnis HUNT, R. D., M. P. ANDERSON u. L. V. CHALIFOUX (1978): Spontaneous infectious diseases of marmosets. Primates Med 10, 239-253 JANG, Y. J., H. J. KWON, H. W. PARK u. B. J. LEE (2006): Detection of rhinovirus in turbinate epithelial cells of chronic sinusitis. Am J Rhinol 20, 634-636 JAQUISH, C. E., T. B. GAGE u. S. D. TARDIF (1991): Reproductive factors affecting survivorship in captive Callitrichidae. Am J Phys Anthropol 84, 291-305 JENSON, H. B., Y. ENCH, Y. ZHANG, S. J. GAO, J. R. ARRAND u. M. MACKETT (2002): Characterization of an Epstein-Barr virus-related gammaherpesvirus from common marmoset (Callithrix jacchus). J Gen Virol 83, 1621-1633 JONES, T. C., U. MOHR u. R. D. HUNT (1993): Monographs on Pathology of Laboratory Animals. Springer Verlag, Berlin JUAN-SALLES, C., A. MARCO u. M. DOMINGO (1998) Intestinal crytococcosis in a common marmoset (Callithrix jacchus) J Med Primatol 27, 298-302 KASPAREIT, J., S. FRIDERICHS-GROMOLL, E. BUSE u. G. HABERMANN (2007): Spontaneous neoplasms observed in cynomolgus monkeys (Macaca fascicularis) during a 15-year period. Exp Toxicol Pathol 59, 163-169 KATSOUYANNI, K. (1995): Health effects of air pollution in southern Europe: are there interacting factors? Environ Health Perspect 103 Suppl 2, 23-27 KEIPER, I. (2002) Qualitative und quantitative bakteriologische und virologische Untersuchungen zur Erhebung des Hygienestatus verschiedener öffentlicher Toilettenanlagen einer südwestdeutschen Großstadt. Dissertation. Institut für Tier- und Umwelthygiene der Tierärztlichen Fakultät der Freien Universität KIM, J. C. u. S. S. KALTER (1975): A review of 105 necropsies in captive baboons (Papio cynocephalus). Lab Anim 9, 233-239 KITANO, M., Y. ITOH, M. KODAMA, H. ISHIGAKI, M. NAKAYAMA, T. NAGATA, H. ISHIDA, H. TSUCHIYA, R. TORII, K. BABA, R. YOSHIDA, A. SATO u. K. OGASAWARA (2010): Establishment of a cynomolgus macaque model of influenza B virus infection. Virology 407, 178-184 98 Literaturverzeichnis KONDGEN, S., S. SCHENK, G. PAULI, C. BOESCH u. F. H. LEENDERTZ (2010): Noninvasive monitoring of respiratory viruses in wild chimpanzees. Ecohealth 7, 332-341 KONDO, H., Y. WADA, G. BANDO, M. KOSUGE, K. YAGI u. Y. OKU (1996): Alveolar hydatidosis in a gorilla and a ring-tailed lemur in Japan. J Vet Med Sci 58, 447-449 KOORIYAMA, T., A. INABA, T. NISHIDA u. T. IWAKI (2010): Case report of helminths and lung mite infection in the red-tailed monkey, Cercopithecus ascanius schmidti, in Mahale Mountains National Park, Tanzania. Primates 51, 183-188 KUIKEN, T., G. F. RIMMELZWAAN, G. VAN AMERONGEN u. A. D. OSTERHAUS (2003): Pathology of human influenza A (H5N1) virus infection in cynomolgus macaques (Macaca fascicularis). Vet Pathol 40, 304-310 KUIKEN, T., B. G. VAN DEN HOOGEN, D. A. VAN RIEL, J. D. LAMAN, G. VAN AMERONGEN, L. SPRONG, R. A. FOUCHIER u. A. D. OSTERHAUS (2004): Experimental human metapneumovirus infection of cynomolgus macaques (Macaca fascicularis) results in virus replication in ciliated epithelial cells and pneumocytes with associated lesions throughout the respiratory tract. Am J Pathol 164, 1893-1900 KUROWSKI, R. u. M. OSTAPCHUK (2002): Overview of histoplasmosis. Am Fam Physician 66, 2247-2252 LE, T. G., L. NGO, S. MEHTA, V. D. DO, T. Q. THACH, X. D. VU, D. T. NGUYEN u. A. COHEN (2012): Effects of short-term exposure to air pollution on hospital admissions of young children for acute lower respiratory infections in Ho Chi Minh City, Vietnam. Res Rep Health Eff Inst 169, 73-83 LEATHERS, C. W. u. T. E. HAMM, JR. (1976): Naturally occurring tuberculosis in a squirrel monkey and a cebus monkey. J Am Vet Med Assoc 169, 909-911 LEE, C. H., L. WILCOX, K. CHORNEYKO u. A. MCIVOR (2008): Coccidioides immitis: two cases of misidentified mycosis. Can Respir J 15, 377-379 LEVER, M. S., A. J. STAGG, M. NELSON, P. PEARCE, D. J. STEVENS, E. A. SCOTT, A. J. SIMPSON u. M. J. FULOP (2008): Experimental respiratory anthrax infection in the common marmoset (Callithrix jacchus). Int J Exp Pathol 89, 171-179 LEVY, B. M. u. R. R. MIRKOVIC (1971): 99 Literaturverzeichnis An epizootic of measles in a marmoset colony. Lab Anim Sci 21, 33-39 LOUIE, J. K., A. E. KAJON, M. HOLODNIY, L. GUARDIA-LABAR, B. LEE, A. M. PETRU, J. K. HACKER u. D. P. SCHNURR (2008): Severe pneumonia due to adenovirus serotype 14: a new respiratory threat? Clin Infect Dis 46, 421-425 LUDLAGE, E. u. K. MANSFIELD (2003): Clinical care and diseases of the common marmoset (Callithrix jacchus). Comp Med 53, 369-382 LUZON, M., C. DE LA FUENTE-LOPEZ, E. MARTINEZ-NEVADO, J. FERNANDEZ-MORAN u. F. PONCEGORDO (2010): Taenia crassiceps cysticercosis in a ring-tailed lemur (Lemur catta). J Zoo Wildl Med 41, 327-330 MACPHAIL, M., J. H. SCHICKLI, R. S. TANG, J. KAUR, C. ROBINSON, R. A. FOUCHIER, A. D. OSTERHAUS, R. R. SPAETE u. A. A. HALLER (2004): Identification of small-animal and primate models for evaluation of vaccine candidates for human metapneumovirus (hMPV) and implications for hMPV vaccine design. J Gen Virol 85, 1655-1663 MATHIS, G. (2007): Bildatlas der Lungen- und Pleurasonographie Springer Medizin Verlag, Berlin, Heidelberg MÄTZ-RENSING, K., J. WINKELMANN, T. BECKER, I. BURCKHARDT, M. VAN DER LINDEN, S. KONDGEN, F. LEENDERTZ u. F. J. KAUP (2009): Outbreak of Streptococcus equi subsp. zooepidemicus infection in a group of rhesus monkeys (Macaca mulatta). J Med Primatol 38, 328-334 MCARTHUR-VAUGHAN, K. u. L. J. GERSHWIN (2002): A rhesus monkey model of respiratory syncytial virus infection. J Med Primatol 31, 61-73 MCCLURE, H. M., J. CHANG, W. KAPLAN u. J. M. BROWN (1976): Pulmonary nocardiosis in an orangutan. J Am Vet Med Assoc 169, 943-945 MCGAVIN, M. D. u. J. F. ZACHARY (2009): Pathologie der Haustiere. Elsevier, London 100 Literaturverzeichnis MILLER, G. F. (1994): Bronchiolar-alveolar adenoma in a rhesus monkey (Macaca mulatta). Vet Pathol 31, 388-390 MILLER, G. F. u. J. W. OWENS (1999): Ultrastructural characterization of the agent of systemic yeast infection of owl monkeys. Med Mycol 37, 139-145 NEUSSER, M., R. STANYON, F. BIGONI, J. WIENBERG u. S. MULLER (2001): Molecular cytotaxonomy of New World monkeys (Platyrrhini) - comparative analysis of five species by multi-color chromosome painting gives evidence for a classification of Callimico goeldii within the family of Callitrichidae. Cytogenet Cell Genet 94, 206-215 O'BRIEN, A. W., L. N. BINN, R. H. HALL, R. J. BEATTIE u. R. H. MARCHWICKI (1981): Measles virus antibodies in a laboratory colony of owl monkeys (Aotus trivirgatus). Lab Anim 15, 343-345 OKOH, A. E. J. u. R. A. OCHOLI (1986): A fatal case of Pasteurellosisi in a Patas monkey in Jos Zoo, Nigeria. J Zoo Anim Med 17, 55-56 PAL, M., G. D. DUBE u. B. S. MEHROTRA (1984): Pulmonary cryptococcosis in a rhesus monkey (Macaca mulatta). Mycosis 27, 309-312 PAZOKI, M., H. MOAZAMI GOODARZI, A. HASHEMI TAHERI, S. SEIFIRAD, N. NEMATOLLAHI u. O. PAKNEJAD (2012): Prevalence of tuberculosis in patients with anthracosis: study on 150 subjects. Arch Iran Med 15, 128-130 PERRYMAN, L. E. (2004): Molecular pathology of severe combined immunodeficiency in mice, horses, and dogs. Vet Pathol 41, 95-100 PHILIPP, M. T., J. E. PURCELL, D. S. MARTIN, W. R. BUCK, G. B. PLAUCHE, E. P. RIBKA, P. DENOEL, P. HERMAND, L. E. LEIVA, G. J. BAGBY u. S. NELSON (2006): Experimental infection of rhesus macaques with Streptococcus pneumoniae: a possible model for vaccine assessment. J Med Primatol 35, 113-122 POTKAY, S. (1992): Diseases of the Callitrichidae: a review. J Med Primatol 21, 189-236 POWER, M. L. u. E. W. MYERS (2009): Digestion in the common marmoset (Callithrix jacchus), a gummivore-frugivore. Am J Primatol 71, 957-963 101 Literaturverzeichnis PUSCH, D., S. IHLE, M. LEBUHN, I. GRAEBER u. J. M. LOPEZ-PILA (2005): Quantitative detection of enteroviruses in activated sludge by cell culture and real-time RT-PCR using paramagnetic capturing. J Water Health 3, 313-324 RAMER, J. C., R. L. GARBER, K. E. STEELE, J. F. BOYSON, C. O'ROURKE u. J. A. THOMSON (2000): Fatal lymphoproliferative disease associated with a novel gammaherpesvirus in a captive population of common marmosets. Comp Med 50, 59-68 REICHE, J. u. B. SCHWEIGER (2009): Genetic variability of group A human respiratory syncytial virus strains circulating in Germany from 1998 to 2007. J Clin Microbiol 47, 1800-1810 RICHARDSON-WYATT, L. S., R. B. BELSHE, W. T. LONDON, D. L. SLY, E. CAMARGO u. R. M. CHANOCK (1981): Respiratory syncytial virus antibodies in nonhuman primates and domestic animals. Lab Anim Sci 31, 413-415 RICHTER, C. B., G. L. HUMASON u. J. H. GODBOLD, JR. (1978): Endemic Pneumocystis carinii in a marmoset colony. J Comp Pathol 88, 171-180 RIMMELZWAAN, G. F., T. KUIKEN, G. VAN AMERONGEN, T. M. BESTEBROER, R. A. FOUCHIER u. A. D. OSTERHAUS (2001): Pathogenesis of influenza A (H5N1) virus infection in a primate model. J Virol 75, 6687-6691 RIVAILLER, P., Y. G. CHO u. F. WANG (2002): Complete genomic sequence of an Epstein-Barr virus-related herpesvirus naturally infecting a new world primate: a defining point in the evolution of oncogenic lymphocryptoviruses. J Virol 76, 12055-12068 ROUSSILHON, C., J. M. POSTAL u. P. RAVISSE (1987): Spontaneous cryptococcosis of a squirrel monkey (Saimiri sciureus) in French Guyana. J Med Primatol 16, 39-47 ROWE, N. u. W. ROWE (1996): Pictorial Guide to the Living Primates Pogonias Press, New York Pogonias Press, New York SANCINI, A., F. TOMEI, G. TOMEI, T. CACIARI, V. DI GIORGIO, J. C. ANDRE, P. PALERMO, G. ANDREOZZI, N. NARDONE, M. P. SCHIFANO, M. FIASCHETTI, C. CETICA u. M. CIARROCCA (2012): Urban pollution. G Ital Med Lav Ergon 34, 187-196 102 Literaturverzeichnis SANTIBANEZ, S., A. HEIDER, E. GERIKE, A. AGAFONOV u. E. SCHREIER (1999): Genotyping of measles virus isolates from central Europe and Russia. J Med Virol 58, 313-320 SCHILDGEN, O., A. SIMON u. J. WILLIAMS (2007): Animal models for human metapneumovirus (HMPV) infections. Vet Res 38, 117-126 SCHILLER, C. A., M. J. WOLFF, L. MUNSON u. R. J. MONTALI (1989): Streptococcus zooepidemicus infections of possible horsemeat source in red-bellied tamarins and Goeldie´s monkeys. J Zoo Wildl Med 5, 322-327 SCHULZE, M., A. NITSCHE, B. SCHWEIGER u. B. BIERE (2010): Diagnostic approach for the differentiation of the pandemic influenza A(H1N1)v virus from recent human influenza viruses by real-time PCR. PLoS One 5, e9966 SEEMA, A., D. K. AGARWAL u. A. DHEERAJ (2010): Pulmonary anthracosis in a monkey: a case report. J Immunol Immunopath 12, 185-192 SHAHDA, S., W. G. CARLOS, P. J. KIEL, B. A. KHAN u. C. A. HAGE (2011): The human metapneumovirus: a case series and review of the literature. Transpl Infect Dis 13, 324-328 SHIMOI, A., C. KAKINUMA, C. KUWAYAMA u. W. MITSUTOSHI (1998): Comparison of spontaneous minor lesions in wild-caught and laboratory-bred monkeys. J Toxical Path 2, 85-94 SHINKI, T., Y. SHIINA, N. TAKAHASHI, Y. TANIOKA, H. KOIZUMI u. T. SUDA (1983): Extremely high circulating levels of 1 alpha,25-dihydroxyvitamin D3 in the marmoset, a new world monkey. Biochem Biophys Res Commun 114, 452-457 SHIRAI, N. u. F. J. GEOLY (2010): Eosinophilic airway inflammation in a cynomolgus monkey. Vet Pathol 47, 318-321 SIMOES, E. A., A. R. HAYWARD, E. M. PONNURAJ, J. P. STRAUMANIS, K. R. STENMARK, H. L. WILSON u. P. G. BABU (1999): Respiratory syncytial virus infects the Bonnet monkey, Macaca radiata. Pediatr Dev Pathol 2, 316-326 103 Literaturverzeichnis SKIADOPOULOS, M. H., S. BIACCHESI, U. J. BUCHHOLZ, J. M. RIGGS, S. R. SURMAN, E. AMAROCARAMBOT, J. M. MCAULIFFE, W. R. ELKINS, M. ST CLAIRE, P. L. COLLINS u. B. R. MURPHY (2004): The two major human metapneumovirus genetic lineages are highly related antigenically, and the fusion (F) protein is a major contributor to this antigenic relatedness. Journal of virology 78, 6927-6937 SMITH, H. A., T. C. JONES u. R. D. HUNT (1978): Veterinary Pathology. Lea & Febiger, Philadelphia SNYDER, S., T. PEACE, O. SOAVE u. J. LUND (1970): Tuberculosis in an owl monkey (Aotus trivirgatus). J Am Vet Med Assoc 157, 712-713 SOLTANI, R., H. KHALILI, A. ABDOLLAHI, M. RASOOLINEJAD u. S. DASHTI-KHAVIDAKI (2012): Nosocomial Gram-positive antimicrobial susceptibility pattern at a referral teaching hospital in Tehran, Iran. Future Microbiol 7, 903-910 SUTHERLAND, S. D., J. D. ALMEIDA, P. S. GARDNER, M. SKARPA u. J. STANTON (1986): Rapid diagnosis and management of parainfluenza I virus infection in common marmosets (Callithrix jacchus). Lab Anim 20, 121-126 SZENTIKS, C. A., S. KONDGEN, S. SILINSKI, S. SPECK u. F. H. LEENDERTZ (2009): Lethal pneumonia in a captive juvenile chimpanzee (Pan troglodytes) due to human-transmitted human respiratory syncytial virus (HRSV) and infection with Streptococcus pneumoniae. J Med Priomatol 38, 236-240 TAPPE, D., K. BREHM, M. FROSCH, A. BLANKENBURG, A. SCHROD, F. J. KAUP u. K. MATZ-RENSING (2007): Echinococcus multilocularis infection of several Old World monkey species in a breeding enclosure. Am J Trop Med Hyg 77, 504-506 TARDIF, S. D., D. A. SMUCNY, D. H. ABBOTT, K. MANSFIELD, N. SCHULTZ-DARKEN u. M. E. YAMAMOTO (2003): Reproduction in captive common marmosets (Callithrix jacchus). Comp Med 53, 364-368 TONG, S., S. W. CHERN, Y. LI, M. A. PALLANSCH u. L. J. ANDERSON (2008): Sensitive and broadly reactive reverse transcription-PCR assays to detect novel paramyxoviruses. J Clin Microbiol 46, 2652-2658 TSUBOTA, K., S. NAKATSUJI, M. MATSUMOTO, S. FUJIHIRA, K. YOSHIZAWA, Y. OKAZAKI, Y. MURAKAMI, A. ANAGAWA, Y. OKU u. Y. OISHI (2009): Abdominal cysticercosis in a cynomolgus monkey. Vet Parasitol 161, 339-341 104 Literaturverzeichnis UMEMURA, T., H. INAGAKI, M. GORYO u. C. ITAKURA (1985): Aspiration pneumonia with adenovirus infection in a Japanese macaque (Macaca fuscata fuscata). Lab Anim 19, 39-41 VAN BINNENDIJK, R. S., R. W. VAN DER HEIJDEN u. A. D. OSTERHAUS (1995): Monkeys in measles research. Curr Top Microbiol Immunol 191, 135-148 WACHTMAN, L. M. u. K. G. MANSFIELD (2008): Opportunistic infections in immunologically compromised nonhuman primates. ILAR J 49, 191-208 WEVERS, D., S. METZGER, F. BABWETEERA, M. BIEBERBACH, C. BOESCH, K. CAMERON, E. COUACYHYMANN, M. CRANFIELD, M. GRAY, L. A. HARRIS, J. HEAD, K. JEFFERY, S. KNAUF, F. LANKESTER, S. A. LEENDERTZ, E. LONSDORF, L. MUGISHA, A. NITSCHE, P. REED, M. ROBBINS, D. A. TRAVIS, Z. ZOMMERS, F. H. LEENDERTZ u. B. EHLERS (2011): Novel adenoviruses in wild primates: a high level of genetic diversity and evidence of zoonotic transmissions. J Virol 85, 10774-10784 WILKINSON, L. M., J. M. WALLACE u. J. M. CLINE (1999): Disseminated blastomycosis in a rhesus monkey (Macaca mulatta). Vet Pathol 36, 460-462 WOLTERS, J u. K. IMMELMANN (1988): Krallenaffen In: Grzimek, B (Hrsg.): Grzimeks Enzyklopädie. Säugetiere. Band 2 Kindler Verlag, München, S. 183-205 XATZIPSALTI, M. u. N. G. PAPADOPOULOS (2007): Cellular and animal models for rhinovirus infection in asthma. Contrib Microbiol 14, 33-41 YAMATE, J., T. IZAWA, M. KUWAMURA, F. MITSUNAGA u. S. NAKAMURA (2009): Vasoformative disorder, resembling littoral cell angioma, of the spleen in a geriatric Japanese macaque (Macaca fuscata). Vet Pathol 46, 520-525 YAMATE, J., A. TOMITA, M. KUWAMURA, F. MITSUNAGA u. S. NAKAMURA (2007): Spontaneous peritoneal malignant mesothelioma in a geriatric japanese macaque (Macaca fuscata). Exp Anim 56, 155-159 YANAI, T., M. A. SIMON, F. D. DODDY, K. G. MANSFIELD, D. PAULEY u. A. A. LACKNER (1999): Nodular Pneumocystis carinii pneumonia in SIV-infected macaques. Vet Pathol 36, 471-474 105 Literaturverzeichnis ZHIVOTOVSKAIA, I. A. (1969): [Blood carbonic anhydrase activity in anthracosis (anthracosilicosis) and pulmonary emphysema]. Sov Med 32, 34-39 ZOU, S., Q. LUO, Z. CHEN, A. CHENG, M. WANG, D. ZHU, R. JIA, F. LIU, X. CHEN, Y. ZHOU, F. BI u. Z. YANG (2010): Isolation, identification of Streptococcus pneumoniae from infected rhesus monkeys and control efficacy. J Med Primatol 39, 417-423 106 Anhang 9 ANHANG 9.1 Göttinger Mischung Ketamin (100 mg/kg) 5 ml Xylazin (10 %ig) 1 ml Atropin (1 %ig) 0,1 ml Aqua ad injectionem 3,9 ml Dosis: 0,2 ml/200 g/KGW; i. m. 9.2 Fixierlösung 10 %iges neutral gepuffertes Formalin: 35 %ige Formaldehydlösung 285 ml Aqua dest. 215 ml Stammlösung A (Phosphatpuffer) 100 ml Stammlösung B (Phosphatpuffer) 400 ml 9.3 Paraffin-Einbettungsprotokoll Wasser (entmineralisiert) für 2 Stunden bei Zimmertemperatur Aufsteigende Alkoholreihe (70 %, 80 %, 96 %, 96 %, 100 %, 100 %) Für jeweils 45 Minuten bzw. 1 Std. bei 35 ºC und Vakuum 3 × Isopropanol für 1 Std. bei 35 ºC bzw. 45 ºC und Vakuum 3 × Paraplast für jeweils 1 Std. bei 62 ºC und Vakuum 9.4 Hämatoxylin -Eosin (HE)-Färbung Die HE-Färbung erfolgte im Färbeautomaten (Varistain Gemini, Fa. Thermo Shandon, Frankfurt am Main) Xylol 1: 5 min. Xylol 2: 2 min. Xylol 3: 2 min. 100 %iges Ethanol: 2 min. 96 %iges Ethanol: 2 min. 107 Anhang 80 %iges Ethanol: 1 min. 70 %iges Ethanol: 1 min. Aqua dest.: 1 min. Zur Kernfärbung Hämalaun nach Mayer (Fa. Merck, Darmstadt): 3 min. Zum Bläuen fließend wässern: 10 min. Gegenfärbung in wässrigem Eosin: 5 min. Fließend wässern: 5 sec. 70 %iges Ethanol: 1 min. 80 %iges Ethanol: 1 min. 96 %iges Ethanol: 1 min. 100 %iges Ethanol: 1 min. 100 %iges Ethanol: 1 min. Xylol: 1 min. Xylol: 3 min. Xylol: 3 min. Eindecken mit Eukitt (Fa. Kindler, Freiburg) 9.5 Berliner-Blau-Reaktion (Eisen-III-Nachweis): Xylol 1: 5 min. Xylol 2: 2 min. Xylol 3: 2 min. 100 %iges Ethanol: 2 min. 96 %iges Ethanol: 2 min. 80 %iges Ethanol: 1 min. 70 %iges Ethanol: 1 min. Aqua dest.: 1 min. Kaliumferrozyanid 10 %: 5 min. 1:1 HCl 20 % und Kaliumferrozyanid 20 %: 30 min. Dreimal in Aqua dest. spülen. Kernechtrot (für Kerne): 5 min. 108 Anhang Dreimal in Aqua dest. spülen Entwässern in aufsteigender Alkoholreihe im Färbeautomaten (Varistain Gemini, Fa. Thermo Shandon, Frankfurt am Main) 70 %iges Ethanol: 1 min. 80 %iges Ethanol: 1 min. 96 %iges Ethanol: 1 min. 100 %iges Ethanol: 1 min. 100 %iges Ethanol: 2 min. Xylol: 1 min. Xylol: 3 min. Xylol: 3 min. Eindecken mit Eukitt (Fa. Kindler, Freiburg) 9.9 Von Kossa-Versilberung Schnitte entparaffinieren, im Färbeautomat bis Aqua dest. 5 %ige Silbernitratlösung in Aqua dest. im Dunkeln: 45 - 60 min. Aqua dest. Fließend: 1 min. In Natriumcarbonat-Formaldehydlösung reduzieren: 2 min. Leitungswasser fließend zum Spülen: 10 min. In 5 %iger Natriumthiosulphatlösung fixieren: 5 min. Leitungswasser fließend zum Spülen: 10 min. Aqua dest. fließend zum Spülen: 1 min. Mit Kernechtrot gegenfärben (für Kerne): 5 min. Aqua dest. fließend zum Spülen: 1 min. Aufsteigende Alkoholreihe im Automaten 3 x Xylol: 1 min. 3 x Xylol: je 2 min. Natriumkarbonat-Formaldehydlösung: 5 g Natriumkarbonat in 25 ml Formaldehyd (35-40 %) und 75 ml Aqua dest. lösen 109 Anhang Kernechtrot: 5 g Aluminiumsulfat in 100 ml Aqua dest. lösen die Aluminiumsulfatlösung erhitzen und 0,1 g Kernechtrot einrühren, bis sich der Farbstoff gelöst hat, erkalten lassen, filtrieren 9.7 Trichromfärbung nach Masson-Goldner Xylol 1: 5 min. Xylol 2: 2 min. Xylol 3: 2 min. 100 %iges Ethanol: 2 min. 96 %iges Ethanol: 2 min. 80 %iges Ethanol: 1 min. 70 %iges Ethanol: 1 min. Aqua dest.: 5 min. Hämatoxylin nach Weigert 2 min. Leitungswasser fließend 10 min. Azophloxin 5 min. Essigsäure 1 %ig 30 sec. Phosphomorlybdänsäure-Orange 2 min. Essigsäure 1 %ig 30 sec. Lichtgrün 5 min. Essigsäure 1 %ig 5 min. Objektträger um die Schnitte herum vorsichtig trocknen Ethanol 96 %ig 2 min. 2 x Ethanol 100 %ig je 2 min. 3 x Xylol je 2 min. Eindecken mit Eukitt (Fa. Kindler, Freiburg) 110 Anhang Hämatoxylin nach Weigert: Stammlösung A: 100 ml Ethanol 96 %ig + 1 g Hämatoxylin (eine Woche reifen lassen) Stammlösung B: 100 ml Aqua dest. + 2,48 g Eisen(III)chlorid + 1 ml konzentrierte HCl Gebrauchslösung: A und B 1:1 vermischen; Gebrauchslösung ist 14 Tage haltbar Gebrauchslösung Azophloxin: Azophloxin 0,5 %ig + 0,2 ml Eisessig in 100 ml Aqua dest. Gebrauchslösung Phosphormolybdänsäure-Orange: 4 g Phosphomolybdänsäure + 2 g Orange G + 100 ml Aqua dest. Gebrauchslösung Lichtgrün: 9.8 Lichtgrün 0,2 %ig + 0,2 ml Eisessig in 100 ml Aqua dest. Amyloid-Färbung mit Kongorot Xylol 1: 5 min. Xylol 2: 2 min. Xylol 3: 2 min. 100 %iges Ethanol: 2 min. 96 %iges Ethanol: 2 min. 80 %iges Ethanol: 1 min. 70 %iges Ethanol: 1 min. Aqua dest.: 5 min. Bei 590 Watt in Mayers Hämalaun bestrahlen: 10 sec. Mit Leitungswasser fließend wässern: 5 min. Bei 590 Watt in Kongorot-Lösung bestrahlen: 50 sec. Kurz in KOH-Lösung differenzieren Mit Aqua dest. fließend wässern 5 min. 111 Anhang Entwässerung in der aufsteigenden Alkoholreihe 70 %iges Ethanol: 1 min. 80 %iges Ethanol: 1 min. 96 %iges Ethanol: 1 min. 100 %iges Ethanol: 1 min. 100 %iges Ethanol: 2 min. Xylol: 1 min. Xylol: 3 min. Xylol: 3 min Eindecken mit Eukitt (Fa. Kindler, Freiburg) Kongorot-Lösung: 1 g Kongorot in 200 ml 50 %igem Ethanol lösen und filtrieren KOH-Lösung: 9.9 2 ml 1 %iges KOH und 200 ml 80 %iges Ethanol mischen Grocott Versilberung Absteigende Alkoholreihe bis Aqua dest. (im Färbeautomat) 5 %ige Chromtrioxidlösung (Chrom (VI)-oxid): 1 Std. Leitungswasser (fließend): 10 min. 1 %ige Natriumdisulfitlösung: 1 min. Leitungswasser (fließend): 5 -10 min. Aqua dest. (fließend): 1 min. Methenamin-Silbernitratgemisch (bei 60 °C): 30 - 60 min. Aqua dest. (fließend): 1 min. 0,2 % ige Goldchloridlösung: 5 min Aqua dest. (fließend): 1 min. 2% ige Natriumthiosulphatlösung: 3 min. Leitungswasser (fließend): 10 min. Lichtgrünlösung: 1 min. Leitungswasser (nicht fließend): 5 min. 112 Anhang Ethanol (96 %): 1 min. 2 x Ethanol (100 %): je 2 min. 3 x Xylol: je 2 min. Methenamin-Silbernitrat Gebrauchslösung: Vor Gebrauch getrennt erwärmen und in folgender Reihenfolge zusammenschütten: 80ml Methenamin 1,5 %ig (1,2 g auf 80ml Aqua dest.) 10ml Silbernitrat 2,5 %ig (0,25 g auf 10ml Aqua dest.) 10ml Natriumtetraborat 1,5 %ig (0,15 g auf 10ml Aqua dest.) Lichtgrün Gebrauchslösung: Lichtgrün 0,2 %ig in 100 ml Aqua dest. lösen, 0,2 ml Eisessig zugeben und filtrieren 9.10 Periodic-Acid-Schiff-Reaktion Entparaffinierung und Rehydratisierung im Färbeautomaten (Varistain Gemini, Fa. Thermo Shandon, Frankfurt am Main): • 0,5 % Perjodsäure: 10 min. • Aqua dest. fließend: 10 min. • Schiff´s Reagenz (Fa. Merck, Darmstadt): 15 min. • Aqua dest.: wenige sec. • Leitungswasser fließend: 10 min. • Gegenfärbung mit Mayers Hämalaun: 30 sec. • Bläuen in Leitungswasser: 10 min. • Entwässerung in der aufsteigenden Alkoholreihe im Färbeautomaten VaristainGemini (Fa. Shandon, Frankfurt am Main): • 70 %iges Ethanol: 1 min. • 80 %iges Ethanol: 1 min. • 96 %iges Ethanol: 1 min. • 100 %iges Ethanol: 1 min. • 100 %iges Ethanol: 2 min. 113 Anhang • Xylol: 1 min. • Xylol: 3 min. • Xylol: 3 min. • Eindecken mit Eukitt (Fa. Kindler, Freiburg) Karbolfuchsinlösung: Fuchsinlösung: 4 g basisches Fuchsin in 20 ml Ethanol 95 % lösen Phenollösung: 8 g Phenolkristalle durch leichtes Erwärmen verflüssigen dann 100 ml Aqua dest. zufügen Zur Herstellung der Karbolfuchsinlösung die Fuchsin- und die Phenollösung mit Magnetrührer über Nacht bei 36 °C mischen. Vor Gebrauch filtrieren. HCl-Alkohol 2: 500ml 70 % Alkohol 1,25 ml konz., rauchende HCl Giemsafärbung Entparaffinierung und Rehydratisierung im Färbeautomaten (Varistain Gemini, Fa. Thermo Shandon, Frankfurt am Main): Giemsalösung für 25 min. Aqua dest. für wenige sec. Differenzieren in 0,5%iger Essigsäure bis die Schnitte einen rötlichen Schimmer bekommen. 90%iger Alkohol für wenige sec. Isopropanol: 5 min. Isopropanol: 5 min. Xylol: 5 min. Eindecken mit Eukitt (Fa. Kindler, Freiburg) 9.11 Anti-Vimentin Immunhistochemie Entparaffinierung Cell Conditioning 114 Anhang 1 Tropfen von Option 1 auftragen, LCS auftragen und für 8 min. inkubieren. Antikörper-Titration Standard AK-Inkubation Inkubieren für 32 min. (primary Antibody) Post Fixation 1 Tropfen von Post-Fixativ auftragen, LCS (liquid coverslip) auftragen und für 8 min. inkubieren. 2ter Antikörper 2ter automatischer Antikörperauftrag 1 Tropfen von Nachweis Nr. 2 (Universal secondary Antibody) auftragen, und für 24 min inkubieren. Gegenfärben 1 Tropfen von Gegenfärbung auftragen, LCS auftragen und für 4 min. inkubieren. 1 Tropfen von Nach-Gegenfärbung auftragen, LCS auftragen und für 4 min. inkubieren. Anti-smooth muscle actin Immunhistochemie Entparaffinierung Cell Conditioning 1 Tropfen von Option 1 auftragen, LCS auftragen und für 8 min. inkubieren. Antikörper-Titration Standard AK-Inkubation Inkubieren für 32 min. (primary Antibody) Post Fixation 1 Tropfen von Post-Fixativ auftragen, LCS auftragen und für 8 min. inkubieren. 2ter Antikörper 2ter automatischer Antikörperauftrag 1 Tropfen von Nachweis Nr. 2 (Universal secondary Antibody) auftragen, und für 24 min. inkubieren. Gegenfärben 115 Anhang 1 Tropfen von Gegenfärbung auftragen, LCS auftragen und für 4 min. inkubieren. 1 Tropfen von Nach-Gegenfärbung auftragen, LCS auftragen und für 4 min. inkubieren 9.12 Lugolsche Lösung Kalium-Iodid 7,5 g in Aqua dest. lösen 18 ml Jod 0,5 g mit Aqua dest. auffüllen 100 ml Durchführung der Färbung: 1 Tropfen Lugolsche Lösung mit etwas Kot verrühren, mit einem Deckglas abdecken und mikroskopieren 9.13 Protokolle für die Bakteriologie Blutagar-Platte Columbia-Agar Basis (Fa. Merck, Darmstadt) 42 g Aqua dest. 1000 ml bei 121 °C 15 min. autoklavieren, auf 50 °C abkühlen lassen Schafblut (Fa. Oxoid, Wesel) 80 ml in Petrischalen abfüllen MacConkey-Agar MacConkey- Agar für die Mikrobiologie (Fa. Merck, Darmstadt) 50 g Aqua dest. 1000 ml zu Platten gießen und bei 121 °C 15 min. autoklavieren Salmonellen-Platte Salmonella-Agar nach ÖNÖZ zur Züchtung von Salmonellen (Fa. Merck, Darmstadt) 80, 5 g Aqua dest. 1000 ml lösen durch Erhitzen und zu Platten gießen Campylobacter-Platte 116 Anhang Campylobacter-Selektivagar (Basis) für die Mikrobiologie (Fa. Merck, Darmstadt) 40 g Aqua dest. 1000 ml bei 121 °C 15 min. autoklavieren; nach Abkühlung auf 45 - 50 °C Campylobacter Selektivsupplement (Fa. Merck, Darmstadt) und 5 - 7 % Blut einmischen Salmonellenanreicherungsbouillon Salmonella-Anreicherungsbouillon nach RAPPAPORT für die Mikrobiologie (Fa. Merck, Darmstadt) 54 g Aqua dest. 1000 ml in Röhrchen abfüllen und 20 min. bei 115 °C autoklavieren 9.14 NucleoSpin RNA II Fa. Macherey-Nagel, Düren Produktnummer: 740955.10 9.15 Reverse Transkriptase Kit Omniscript, Fa. Qiagen, Hilden Invitrogen Produktnummer: 100000840 Konzentration 40 units/µl Master mix (MM): 10x Buffer RT 2 μl dNTP Mix (5 mM je dNTP) 2 μl Oligo-dT primer (80 pmol) RNase Inhibitor (40 units/μl) 0,25 μl Omniscript Reverse Transcriptase (4 Einheiten pro μl) 1 µl RNase-freies Wasser Template RNA Template RNA; 500 ng (per 20 μl Reaktion) Total volume: 20 μl 117 Anhang 9.16 Nested PCR Master mix äußere Primer: 1 µl forward primer 1 µl reverse primer 2 µl 10fach Puffer 0,15 µl dNTP´s 0,2 µl taq-Polymerase 10,65 µl Wasser 15 µl MM + 5,0 µl cDNA Master mix innere Primer: 1 µl forward primer 1 µl reverse primer 2 µl 10fach Puffer 0,15 µl dNTP´s 0,2 µl taq-Polymerase 13,15 µl Wasser 17,5 µl MM + 2,5 µl PCR-Produkt 118 Anhang Abb. 19: Chronizität der Entzündungszellinfiltrate. akut 6% akut-subakut 8% subakut 86% Abb. 20: Verteilungsmuster der Entzündungszellinfiltrate. 40 37 35 30 Anzahl 25 20 16 15 10 6 5 5 2 0 fokal oligofokal multifokal Verteilungsmuster 119 multifokalkonfluierend diffus Anhang Abb. 21: Schweregrade der interstitiellen Pneumonien. mittelhochgradig 2% hochgradig 1% mittelgradig 20% geringmittelgradig 13% geringgradig 64% Tab. 11 : Häufigkeit des Auftretens von Bronchopneumonien im Tierbestand des DPZ. Art der Grad der BronchoEntzündung pneumonie Sektionsnummer Geschlecht Alter der Tiere Ausprägung Alter der Veränderung 4643 weiblich juvenil purulent hochgradig diffus akut-subakut 4880 weiblich adult purulent hochgradig diffus akut-subakut 5936 weiblich adult purulent hochgradig multifokal akut 6450 weiblich neugeboren purulent hochgradig multifokalkonfluierend subakut 6478 männlich neugeboren purulent hochgradig fokal subakut 7733 männlich juvenil purulent hochgradig multifokalkonfluierend subakut 120 Anhang Tab. 12: Häufigkeit des Auftretens von Lobärpneumonien im Tierbestand des DPZ. Sektionsnummer Geschlecht Alter der Tiere Art der Lobärpneumonie Grad der Entzündung Ausprägung Alter der Veränderung 5317 weiblich neugeboren fibrinös gering- bis mittelgradig multifokal subakut 5325 männlich neugeboren fibrinös mittelgradig multifokalkonfluierend akut-subakut hochgradig multifokalkonfluierend subakut 6310 weiblich adult Pleuropneumonie, fibröspurulent 6382 weiblich juvenil purulent hochgradig multifokalkonfluierend subakut 7813 weiblich adult purulent mittelgradig multifokalkonfluierend akut-subakut 8120 männlich adult fibröspurulent hochgradig fokal akut 8121 männlich adult purulent mittelgradig multifokalkonfluierend akut 8198 weiblich juvenil fibröspurulent geringgradig multifokal akut adult Pleuropneumonie, fibröspurulent; mittelgradig multifokal chronischaktiv 8396 weiblich Tab. 13: Häufigkeit des Auftretens von Alveolitiden im Tierbestand des DPZ. Sektionsnummer Geschlecht Alter der Tiere Grad der Entzündung Ausprägung Alter der Veränderung 6476 männlich juvenil geringgradig multifokal akut 8093 weiblich juvenil mittelgradig multifokalkonfluierend subakut 8386 männlich adult geringgradig multifokal subakut 121 Anhang Tab. 14: Häufigkeit des Auftretens von bronchointerstitiellen Pneumonien im Tierbestand des DPZ. Sektionsnummer Geschlecht Alter der Tiere Grad der Entzündung Ausprägung Alter der Veränderung 7857 männlich adult hochgradig diffus subakut 7286 weiblich juvenil mittelgradig multifokal akut Tab. 15: Weißbüschelaffe aus dem Tierbestand des DPZ mit einer granulomatösen Pneumonie. Sektionsnummer Geschlecht Alter des Tieres Grad der Entzündung Ausprägung Alter der Veränderung 7165 weiblich adult geringgradig fokal chronisch Tab. 16: Häufigkeit des Auftretens von Lungentumoren im Tierbestand des DPZ. Sektionsnummer Geschlecht Alter des Tieres Art des Tumors 6188 männlich adult malignes BZelllymphom 6758 weiblich juvenil malignes BZelllymphom 6923 weiblich juvenil Fibrosarkom 6944 männlich adult malignes T-zellreiches B-Zell Lymphom 8428 weiblich adult Lymphom 122 Anhang Tab. 17: Geschlechterverhältnis der Tiere mit minimalen Zellinfiltraten in den alveolären Septen. Weibchen Männchen keine Angabe Gesamt 287 285 1 573 Tiere mit minimalen Entzündungszellinfiltraten 34 32 0 66 Gesamt 321 317 1 639 Tiere ohne minimale Entzündungszellinfiltrate Tab. 18: Geschlechterverhältnis der Tiere mit interstitiellen Pneumonien. Weibchen Männchen keine Angabe Gesamt Tiere ohne interstitielle Pneumonie 217 216 1 434 Tiere mit interstitieller Pneumonie 104 101 0 205 Gesamt 321 317 1 639 Tab. 19: Häufigkeit des Auftretens interstitieller Pneumonien in den verschiedenen Altersgruppen. Neugeboren Juvenil Adult Gesamt Tiere ohne interstitielle Pneumonie 119 81 234 434 Tiere mit interstitieller Pneumonie 51 60 94 205 Gesamt 170 141 328 639 123 Anhang Tab. 20: Hauptbefunde der Tiere mit Masernnachweis. Sektionsnummer Alter (Monate) 8506 34 8524 44 Geschlecht weiblich weiblich Befund: Lunge Befund: Dünndarm Befund: Dickdarm o.b.B. hochgradige, chronische, lymphozytäre Enteritis mittelgradige, lymphozytäre Colitis mittel- hochgradige, chronisch-aktive, erosive Duodenitis o.b.B. minimale Alveolarhistiozytose sowie multifokal, geringgradige, gemischtzellige Leukozytostase in den Alveolarsepten; kleine Mineralisationsherde Tab. 21: Sequenzbereich des PCR-Produktes der beiden Tiere mit positivem Masernbefund Sektionsnummer Sequenzbereich 8506 5’TTGGGTCGGCTCGCAAAGGCGGTTACGGCCCCAGACACGGCAGCTGATTCGGA GCTAAGAMGGTGGATAAAGTACACCCAACAAAGAAGGGTGGTTGGTGAATTTAG GTTGGARAGAAAATGGTTGGATGTGGTGAGAAACAGGATTGCCGAGGACCCTCC TTACGCCGATTCATGGTCGCTCTAATCCTGGATATCAAGAGAACACCCGGGAACA AACCCAGGATTGCTGAAATGATATGTGACATTGATACATATATAGTAGAGGCAGG ATTAGCCAGTTTTATCCTAACTATTAAGTTTGGGATAGAAAC -3‘ 8524 5‘MTCTYGCTCGCAAAGGCGGTTACGGCCCCAGACACGGCAGCTGATTCGGAGCTA AGAAGGTGGATAAAGTACACCCAACAAAGAAGGGTGGTTGGTGAATTTAGGTTG GAGAGAAAATGGTTGGATGTGGTGAGAAACAGGATTGCCGAGGACCTCCCTTAC GCCGATTCATGGTCGCTCTAATCCTGGATATCAAGAGAACACCCGGGAACAAACC CAGGATTGCTGAAATGATATGTGACATTGATACATATATGAGAGGCAGGATTAGC CAGTTTTATCCTAACTATTAAGTTTGGGATAGAAACTATGTAT -3‘ 124 Anhang Tab. 22: Immunhistochemische Untersuchung auf B- und T-Lymphozyten in ausgewählten Lungen. Sektionsnummer Geschlecht Alter (Monate) Morphologische Diagnose T-Lymphozyten B-Lymphozyten Makrophagen 8124 weiblich 18 geringgradige, subakute, multifokal-konfluierende interstitielle Pneumonie 448 4 824 7681 männlich 4 geringgradige, subakute, multifokal-konfluierende interstitielle Pneumonie 1121 7 1192 7789 weiblich 8 mittelgradige, subakute, multifokal-konfluierende interstitielle Pneumonie 2515 5 1036 7824 männlich 5 mittelgradige, subakute, multifokal-konfluierende interstitielle Pneumonie 1390 37 602 Tab. 23: Häufigkeit des Auftretens interstitieller Pneumonien in den verschiedenen Geschlechtsgruppen in der Kolonie der externen Labortiereinrichtung. Tiere ohne interstitielle Pneumonie Tiere mit interstitieller Pneumonie Gesamt Weibchen 5 7 12 Männchen 11 11 22 Gesamt 16 18 34 125 Anhang Tab. 24: Häufigkeit des Auftretens interstitieller Pneumonien in den verschiedenen Altersgruppen in der Kolonie der externen Labortiereinrichtung. Tiere ohne interstitielle Pneumonie Tiere mit interstitieller Pneumonie Gesamt Neugeboren 0 2 2 Juvenil 11 10 21 Adult 5 6 11 Gesamt 16 18 34 126 Danksagung Mein besonderer Dank gilt meinem Doktorvater, Herrn Univ.-Prof. Dr. Franz-Josef Kaup, dessen uneingeschränkte Unterstützung mein Interesse und meine Freude an dieser Arbeit beflügelt und ganz wesentlich zu deren Gelingen beigetragen haben. Frau Dr. Martina Bleyer danke ich, dass sie trotz vollen Terminkalenders jederzeit für mich ansprechbar war und mit professionellem Rat und konstruktiven Vorschlägen meine Arbeit begleitete. Frau Dr. Kerstin Mätz-Rensing danke ich für die wertvollen Hilfestellungen, ihre immer freundliche und fachliche Unterstützung sowie die stete Bereitschaft, ihr pathologisches Wissen mit mir zu teilen. An unsere gemeinsamen Sektionen erinnere ich mich mit Freude. Frau Dr. Eva Gruber-Dujardin danke ich für die vielen Anregungen, Ratschläge und Hilfestellungen sowie ihre Unterweisungen auf dem Gebiet der Histopathologie. Herrn Hans-Joachim Helms, Herrn David Ellenberger und den Kollegen aus der Abteilung Medizinische Statistik der Universitätsmedizin Göttingen danke ich für die Unterstützung bei der statistischen Auswertung meiner Daten. Für die freundliche, fachkundige Unterstützung und die enorme Hilfsbereitschaft danke ich dem Assistententeam Nadine Schminke, Andreas Kues und Larissa Hummel. Auch wenn „Hinter der Schleuse“ nicht immer Einigkeit über Tischhöhe und Radiosender bestand, habe ich die Zusammenarbeit mit Herrn Wolfgang Henkel jederzeit genossen und denke mit Freude an viele interessante Sektionen mit ihm zurück. Mein Dank geht ebenso an Frau Elke Lischka und Frau Stefanie Wienstroth, ohne die das „Schleusenteam“ nicht komplett wäre. Frau Tamara Becker danke ich, dass ich sie bei ihren Visiten begleiten durfte, und sie mir lehrreiche und interessante Einblicke in die praktische tierärztliche Tätigkeit ermöglichte. Bei Frau Annette Husung bedanke ich mich herzlich für die Bereitstellung der Informationen zur Primatenhaltung. 127 Meinen aktuellen und ehemaligen im Arbeitskollegen „Doktorandenzimmer“ Christoph Curths, Anne Schmitt, Karen Lampe, Hans-Dieter Lauenstein, Rebecca Hoffmann und Julia Marie Winkelmann danke ich für das gute Arbeitsklima und die vielen witzigen und skurrilen Momente in diesem eigenen kleinen Mikrokosmos. Dank sage ich auch allen nicht namentlich genannten Kollegen aus der Abteilung Infektionspathologie und Primatenhaltung. Es war schön, mit Euch zusammenzuarbeiten. Ein „special thanks“ geht an die Doktoranden der Abteilung Klinische Neurobiologie, mit denen ich gemeinsame Mensastunden, „Cafe-pausitas“ und beachvolleyballlastige Feierabende und Wochenenden verbracht habe. Ohne Euch, Carolina Araya-Callis, Nicole Yee, Enrique Garea-Rodriguez und Adema Ribic, wäre Göttingen für mich nicht Göttingen geworden. Und schließlich empfinde ich besondere Dankbarkeit gegenüber meiner Familie. Vor allem meinen Eltern danke ich von Herzen für ihren bedingungslosen Rückhalt, den sie mir gewährt haben. 128