Versuchsprotokoll: Leitenzyme Aufschluss von Lebergewebe
Werbung

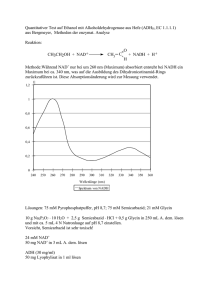
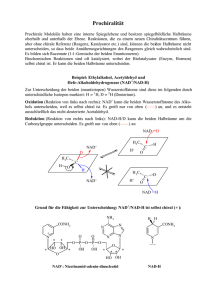
Versuchsprotokoll: Leitenzyme Aufschluss von Lebergewebe / Fraktionierte Zentrifugation / Aufschluss von Mitochondrien / Nachweis von Leitenzymen im Cytosol und Mitochondrienplasma 1.1. Einleitung Für die Trennung der Organellen und des Cytosols benutzt man die Fraktionierte Zentrifugation. Vorher wird das Lebergewebe zerkleinert und dann homogenisiert. Durch die Zentrifugation mit immer steigender Rotationsgeschwindigkeit werden die durch das Homogenisieren freigesetzten Organellen bzw. Plasma voneinander getrennt. Durch die Nachweise von sogenannten Leitenzymen kann man erstens nachweisen, ob die gewonnenen Fraktionen frei von anderen Bestandteilen sind und zweitens, um welche Organellen bzw. um welches Plasma es sich handelt. Daher benutzt man zum Nachweis nur Enzyme, die in einem Kompartiment vorkommen, in einem anderen nicht (hier die Glutamat-Dehydrogenase und die Lactat-Dehydrogenase). Allerdings kann man zur Probe auch ein Enzym benutzten, dass in beiden Kompartimenten vorkommt (hier: die NADP- abhängige Isocitrat-Dehydrogenase). Die gesamte Trennung wird unter ständiger Kühlung durchgeführt, um Abbauprozesse zu verhindern. 1.2. Material und Methode Die in diesem Versuchsteil benutzten Materialien sind im Skript auf den Seiten 20 und 21 angegeben. Die Methode der Zentrifugation ist in folgender Abbildung dargestellt: Abb. 1: Fraktionierte Zentrifugation, aus Skript Seite 22 Seite 1 von 7 Bei der sich daran anschließenden Photometrie wird ausschließlich die Extinktion von NADH bzw. NADPH gemessen. NAD(P)H besitzt Absorptionsmaxima bei 260nm und bei 340nm. NAD(P) dagegen weißt bei 340nm kein Absorptionsmaximum auf, da die Mesomerie im Pyrimidinring aufgehoben wurde. Wird nun NAD(P)H verbraucht, sinkt die Extinktion. Da das NAD(P)H bzw. NAD(P) bei allen drei Reaktionen als Coenzym oxidiert bzw. reduziert wird, macht die Photometrie bei ca. 340nm eine quantitative Aussage über die Enzymkonzentrationen im jeweiligen Plasma. O O H H H C C NH2 NH2 N N R R 2 [H] NAD(P)H + H* NAD(P)+ Extinktion bei 340 nm keine Extinktion bei 340 nm Abb.2: Reaktive Gruppel von NAD(P) 1.3. Versuchsdurchführung Siehe Skript Seiten 20 und 21 1.4. Versuchsergebnisse 1.4.1 Glutamat-DH-Test Mitochondrienplasma (0,02ml) Zeit in s Extinktion 20 0,8 40 0,69 60 0,6 80 100 0,48 0,385 120 140 0,3 0,225 260 280 320 0,035 0,025 300 160 180 200 220 240 0,17 0,125 0,09 0,065 0,05 340 0,02 0,016 0,014 Cytoplasma (0,02ml) Zeit in s Extinktion 30 60 90 1,065 1,065 1,005 120 150 180 210 240 1 0,995 0,995 0,995 0,995 Seite 2 von 7 Glutamat-DH-Test 1,2 1 Extinktion 0,8 0,6 0,4 0,2 0 0 50 100 150 200 250 300 350 400 Zeit in s Mitochondrienplasma Cytoplasma 1.4.2. Isocitrat-DH-Test Mitochondrienplasma (0,8ml) Zeit in s Extinktion 30 60 90 120 150 180 210 240 270 300 0,12 0,215 0,315 0,395 0,47 0,52 0,545 0,55 0,555 0,56 330 360 390 420 0,56 0,56 0,56 0,56 Cytoplasma (0,8ml) Zeit in s Extinktion 30 0,14 60 90 0,23 0,325 120 150 180 0,4 0,47 0,52 210 240 0,55 0,58 270 300 330 360 390 420 0,6 0,61 0,615 0,62 0,62 0,62 Isocitrat_DH-Test 0,7 0,6 Extinktion 0,5 0,4 0,3 0,2 0,1 0 0 50 100 150 200 250 300 350 400 450 Zeit in s Mitochondrienplasma Cytoplasma Seite 3 von 7 1.4.3. Lactat-DH-Test Mitochondrienplasma (0,05ml) Zeit in s Extinktion 30 60 90 120 150 180 210 240 270 300 330 360 390 420 0,74 0,7 0,68 0,65 0,62 0,6 0,57 0,54 0,52 0,5 0,47 0,445 0,42 0,4 Cytoplasma (0,05ml) Zeit in s Extinktion 30 60 90 120 150 180 210 240 270 300 330 0,95 0,7 0,53 0,38 0,23 0,1 0,08 0,04 0,02 0 0 360 390 0 0 Lactat-DH-Test 1 0,9 0,8 Extinktion 0,7 0,6 0,5 0,4 0,3 0,2 0,1 0 0 50 100 150 200 250 300 350 400 450 Zeit in s Mitochondrienplasma Cytoplasma Die Extinktionswerte des Cytoplasmas wurden angehoben, um eine negative Extinktion zu umgehen. Um die Werte mit dem Mitochondrienplasma zu vergleichen, muss der Wert von 0,48 abgezogen werden. Da es aber bei dem Versuch nicht auf den unmittelbaren Extinktionsvergleich ankommt, sonder eher auf den Verlauf der Extinktion, ist diese Manipulation vollkommen legitim. 1.5. Auswertung / Diskussion 1.5.1. Glutamat-DH-Test Die Glutamat-Dehydrogenase (GDH) ist ein weitgehend spezifisches Leberenzym. Die GDH katalysiert folgende Reaktion: Glutamat entsteht in den Mitochondrien bei einer Nebenreaktion des Citratzykluses aus alpha-Ketoglutarat und NH4+: Seite 4 von 7 Glutamat-DH + + alpha-Ketoglutarat + NH4 + NAD(P)H + H Glutamat + H2O + NAD(P)+ Das NADH + H+ ist, wie eingangs schon erwähnt, das Coenzym und wird hier oxidiert. Bei Leberschäden kann das Enzym auch im Blut festgestellt werden und weist somit auf eine Erkrankung hin. Durch die Abnahme der NADH + H+ -Konzentration sinkt die Extinktion im Laufe der Versuchszeit wegen der Umsetzung in NAD+ ab. Dies liegt an der, unter Punkt 1.2 (Seite 2) bereits beschrieben, veränderten Mesomerie des Pyrimidinrings des Nicotinamids. Die Abnahme der Konzentration erfolgt jedoch nur bei der Zugabe von Mitochondrienplasma. Somit liegt eine fast saubere Trennung vom Cytoplasma vor, welches keine enzymatische Aktivität aufweist. Da der Citratzyklus ausschließlich in den Mitochondrien lokalisiert ist und somit auch die Umsetzung von alpha-Ketoglutarat in Glutamat stimmen diese Beobachtung und das erwartete Ergebnis überein. 1.5.2. Isocitrat-DH-Test Die NADP-abhängige Isocitrat-Dehydrogenase (NADP-ICD) kommt in beiden Kompartimenten vor. Dies ist eindeutig an den fast gleichen Extinktionskurven abzulesen. Die Isocitrat-Dehydrogenase (IDH) katatlysiert folgende Reaktion: Isocitrat + NADP+ NADP-ICD alpha-Ketoglutarat + CO2 + NADPH + H+ (Abb. 3, 1.5.2.) „Die IDH ist entweder NAD oder NADP abhängig. In Eukaryonten gibt es mindestens drei Isoformen der IDH: zwei befinden sich in der Mitochondrialen Matrix (eine ist NAD abhängig, die andere NADP abhängig), während die dritte (auch NADP abhänging) im Cytoplasma lokalisiert ist.“ (EMBL EBI, European Bioinformatics Institut, http://www.ebi.ac.uk/interpro/IEntry?ac=IPR001804 ) Somit ist verständlich, dass auch im Cytoplasma eine NADP-ICDH-Funktion festzustellen ist. Die Aufgabe, der NADP-ICD in dem Cytoplasma ist einmal die oben aufgezeigte Reaktion zu katalysieren und somit zweitens auch eine NADPH+H+ -Quelle darzustellen, besonders dann, wenn eine Glucose-6-phosphat dehydrogenase deficiency vorliegt. (Geisbrecht BV, Gould SJ. (1999): The human PICD gene encodes a cytoplasmic and peroxisomal NADP+-dependent isocitrate dehydrogenase. – J Biol Chem.274 (43): 30527-33.) Auch in Peroxisomen ist die NADP-ICD anzutreffen. Da bei der Fraktionierten Zentrifugation Mitochondrien und Peroxisomen gemeinsam „geknackt“ wurden, kann man somit auch annehmen, dass die NADP-ICD aus den Peroxisomen stammt. Dort hat sie die Aufgabe durch die oben bereits gezeigte Reaktion (Abb. 1,1.5.2.) die NADPHKonzentration für intrazelluläre Reduktionen zu regenerieren. (Geisbrecht BV, Gould SJ. (1999): The Seite 5 von 7 human PICD gene encodes a cytoplasmic and peroxisomal NADP+-dependent isocitrate dehydrogenase. – J Biol Chem.274 (43): 30527-33.) In anderen Geweben, wie besonders Muskel und Herz, wo es keine großen NADPH – Konsumenten gibt, hat die NADP-ICD in der Mitochondrienmatrix wahrscheinlich die Aufgabe zusammen mit der H+-Transhydrogenase die Aktivität des Tricarbonsäurezykluses in Abhängigkeit der Energienachfrage zu regulieren. Diese beiden Enzyme regulieren sozusagen die NAD-ICD. Die NAD-ICD treibt den Tricarbonsäurezyklus an, während die NADP-ICD ihn verlangsamt. Die NADP-ICD erhöht den NADPH-Spiegel (in Abhängigkeit der Aktivität der H+-Transhydrogenase). NADPH ist genauso wie ATP ein allosterischer Effektor der NAD-ICD. (Sazanov LA, Jackson JB. (1994): Proton-translocation transhydrogenase and NAD- and NADP-linked isocitrate dehydrogenases operate in a substrate cycle which contributes to fine regulation of the tricarboxylic acid cycle activity in mitochondria. – FEBS Lett: 344 (2-3): 109-16.) Somit wurde die Funktion der NADP-abhängigen ICD in der Mitochondrienmatrix aufgezeigt. 1.5.3. Lactat-DH-Test Die Lactat-Dehydrogenase (LDH) katalysiert die Umsetzung von Pyruvat zu Lactat. Dabei ist die Entstehung des Lactats nicht entscheidend, sondern die Regeneration des NAD+. Die Regeneration des NAD+ erfolgt jedoch nur im anaeroben Stoffwechsel oder bei einer Sauerstoffunterversorgung im Skelettmuskel über die LDH, da NADH + H+ ansonsten in der Atmungskette regeneriert wird. Cytoplasma Lactat-DH Pyruvat Lactat NADH + H+ NAD+ 5. Schritt d. Glycerinaldehyd-3-phosphat 1,3-Bisphosphoglycerat Glycolyse GAP-DH Das NADH + H+ ist, wie eingangs schon erwähnt, das Coenzym und wird hier oxidiert. Durch die Abnahme der NADH + H+ -Konzentration sinkt die Extinktion im Laufe der Versuchszeit wegen der Umsetzung in NAD+ ab. Dies liegt an der, unter Punkt 1.2 (Seite 2) bereits beschrieben, veränderten Mesomerie des Pyrimidinrings des Nicotinamids. Die Abnahme der Konzentration erfolgt jedoch maßgeblich nur bei der Zugabe von Cytoplasmalösung, was auch der Theorie entspricht, da die Glycolyse nur in dem Cytoplasma abläuft. Seite 6 von 7 Die Abnahme bei Zugabe von Mitochondrienplasma kann durch eine Verunreinigung hervorgerufen worden sein, bzw. eine nicht einwandfreie Trennung von Cytosol und Mitochondrienplasma. Allerdings ist die Abnahme der Extinktion sehr flach und im Gegensatz zu der Cytoplasmazugabe gering. Da die Extinktion von NADH + H+ gemessen wird, könnte es sein, dass ein anderes Enzym, welches in der Mitochondrienplasma vorkommt das NADH + H+ oxidiert und somit eine Abnahme der Extinktion zustande kommt. Allerdings lässt sich dies hier nicht nachweisen. Seite 7 von 7