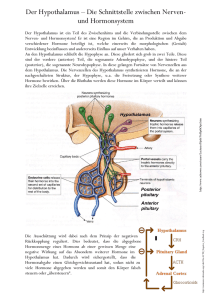
Archivobjekt öffnen
Werbung

Aus dem Institut für Neuroendokrinologie der Universität zu Lübeck Direktor: Prof. Dr. J. Born Die regulatorische Rolle von Cortisol und Wachstumshormon auf die Zytokinproduktion humaner T-Lymphozyten und die Th1/Th2-Balance Inauguraldissertation zur Erlangung des akademischen Grades Doktor der Medizin (Dr. med.) der Medizinischen Fakultät der Universität zu Lübeck Vorgelegt von: Melanie Wolf aus Neubrandenburg Lübeck, 2012 1. Berichterstatter: Professor Dr. med. J. Born 2. Berichterstatter: Priv. - Doz. Dr. med. Kai Mortensen Tag der mündlichen Prüfung: 01.11.2012 Zum Druck genehmigt. Lübeck, den 01.11.2012 -Promotionskommission der Sektion Medizin- 2 Diese Arbeit widme ich meiner Familie 3 Inhaltsverzeichnis Inhaltsverzeichnis Seite Abkürzungsverzeichnis 6 1 Einleitung 8 1.1 Das Immunsystem 8 1.1.1 Zellen des Immunsystems 8 1.1.2 Th1/Th2-Immunbalance 12 1.2 Endokrinum und das Immunsystem 14 1.2.1 Das Glukokortikoid Cortisol 16 1.2.2 Einfluss von Cortisol auf das Immunsystem und die Th1/Th2-Balance 17 1.2.3 Das humane Wachstumshormon GH 18 1.2.4 Einfluss von GH auf das Immunsystem und die Th1/Th2-Balance 19 1.3 Schlaf, Endokrinum und das Immunsystem 19 1.4 Fragestellung und Ziel der Arbeit 24 2 Material und Methoden 25 2.1 Probanden 25 2.2 Reagenzien 25 2.3 Versuchsablauf 26 2.3.1 Zellstimulation und Aktivierung 26 2.3.2 Durchflusszytometrische Analyse 27 2.3.3 Hormonbestimmung 28 2.3.4 Statistische Auswertung der Ergebnisse 29 3 Ergebnisse 30 3.1 Plasma - Hormon - Konzentration innerhalb der Kontrollgruppe 30 4 Inhaltsverzeichnis 3.2 Auswirkungen auf IFN-γ-produzierende CD4-positive T-Helferzellen 31 3.3 Auswirkungen auf IL-4-produzierende CD4-positive T-Helferzellen 33 3.4 Auswirkungen auf das IFN-γ-/ IL-4-Gleichgewicht (Th1/Th2-Balance) 35 4 Diskussion 37 5 Zusammenfassung 54 6 Literaturverzeichnis 56 7 Anhang 82 8 Danksagung 84 9 Lebenslauf 85 10 Veröffentlichung der Arbeitsergebnisse 86 5 Abkürzungsverzeichnis Abkürzungsverzeichnis ACTH: Adrenocorticotropes Hormon (Adrenocorticotropin) APC: Allophycocyanin APZ: Antigenpräsentierende Zellen BFA: Brefeldin A CD: Cluster of Differentiation CRH: Corticoliberin (corticotropin-releasing hormone) CTL: Cytotoxische T-Lymphozyten (cytotoxic T-cells) DC: Dendritische Zellen (dendritic cells) DEX: Dexamethason DFG: Deutsche Forschungsgemeinschaft DMSO: Dimethylsulfoxid FACS: Durchflusszytometer (Fluorescence Activated Cell Sorter) FITC: Fluorescein-Iso-Thio-Cyanat (Fluorochrom) FSC: Vorwärtsstreuung (Frontscatter) GK: Glukokortikoid hGh: Wachstumshormon, somatotrophes Hormon (human growth hormone) HHN – Achse: Hypothalamus-Hypophysen-Nebennierenrinden-Achse HVL: Hypophysenvorderlappen IFN: Interferon Ig: Immunglobulin IGF: Insulinähnlicher Wachstumsfaktor (Insuline-like growth factor) IL: Interleukin MHC: Haupthistokompatibilitätskomplex (Major Histocompatibility Complex) MK: Mineralokortikoid NK-Zellen: Natürliche Killerzellen NNR: Nebennierenrinde nTreg: natürliche regulatorische T-Zellen (natural regulatory T cells) 6 Abkürzungsverzeichnis SEM: Standard Error of the Mean SSC: Seitwärtsstreuung (Sidescatter) SWS: langsamwellige Schlafphase (Slow Wave Sleep) PE: Phycoerythrin PerCP: Peridin Chlorophyll Protein PBMC: mononukleäre Zellen des peripheren Blutes (peripher blood mononuclear cells) PBS: Phosphatgepufferte Salzlösung (phosphate buffered saline) PMA: Phorbol 12-myristate 13-acetate R-A-A-S: Renin-Aldosteron-Angiotensin-System REM: Rapid eye movement Rpm: Umdrehungen pro Minute (revolutions per minute) Th-Zellen: T-Helferzellen TCR: T-Zell Rezeptor (t-cell receptor) TSH: Thyreotropin, thyreotropes Hormon (thyroid stimulating hormone) 7 Einleitung 1. Einleitung Die vorliegende Arbeit beschäftigt sich mit dem Einfluss des Wachstumshormons (GH) und des Cortisols auf das Immunsystem. Insbesondere wird dabei das Verhältnis der Th1- und Th2-Zytokine unter in vitro Bedingungen untersucht. Es galt die Hypothese zu überprüfen, ob niedrige Cortisol- und / oder hohe GHKonzentrationen zu einer Dominanz des Th1-Zytokinprofils führen. Um einen Überblick über die klinische Relevanz des Versuchsvorhabens zu geben, wird im Folgenden auf das Immunsystem und dessen Bestandteile eingegangen. 1.1 Das Immunsystem Der Mensch benötigt ein gut funktionierendes Immunsystem, um in einer Umwelt verschiedenster gesundheitlicher Herausforderungen überleben zu können. In den Organismus eingedrungene körperfremde Strukturen müssen aufgesucht und bekämpft werden. Ebenfalls müssen fehlerhaft veränderte, mutierte (z. B. maligne entartete) körpereigene Zellen gefunden und, bevor Schäden im Organismus auftreten, eliminiert werden. Man unterscheidet eine unspezifische, bereits bei wirbellosen Lebewesen vorhandene, angeborene Immunabwehr von einer adaptiven Immunabwehr. Letztere bietet einen spezifischen Schutz gegen Erreger und ist in der Lage, ein immunologisches Gedächtnis gegen wiederkehrende Pathogene auszubilden. Auf dieses immunologische Gedächtnis stützt sich das Prinzip der Impfung (Janeway et al., 2009; Müller et al., 2008). 1.1.1 Zellen des Immunsystems Die „erste“ Linie der Verteidigung gegen Pathogene wird durch das angeborene Immunsystem realisiert. Hierfür stehen Makrophagen, natürliche Killerzellen (NKZellen), dendritische Zellen (DC), neutrophile, basophile und eosinophile Granulozyten zur Verfügung (Figdor et al., 2002; Houssaint und Sai 1988; Svedmyr und Jondal 1975). Durch Makrophagen werden Pathogene anhand der auf ihrer Oberfläche befindlichen Proteine erkannt, durch Phagozytose zersetzt 8 Einleitung und die so entstandenen Antigene auf der Oberfläche der Makrophagen präsentiert (Aderem und Underhill 1999). Diese Antigenpräsentation führt zur Aktivierung von Zellen der erworbenen Immunabwehr. Die aktivierten Makrophagen leiten durch Freisetzung verschiedener chemischer Botenstoffe, den Zytokinen, lokale Entzündungsreaktionen ein (Kayser et al., 2005). Die 1973 von Ralf Steinmann entdeckten dendritischen Zellen (DC) gehören analog den Makrophagen zu den antigenpräsentierenden Zellen (APZ). Sie vermitteln über aktivierte B- und T-Lymphozyten zwischen angeborener und erworbener Immunabwehr (Banchereau et al., 2000; Steinman und Hemmi 2006; Villadangos und Schnorrer 2007). Natürliche Killerzellen (NK) sind Bestandteil der angeborenen Immunabwehr. Sie sind in der Lage virus-infizierte und entartete Zellen zu erkennen. NK werden über ein komplexes Rezeptorsystem aktiviert und lösen durch Ausschüttung verschiedener Zytotoxine die Apoptose der Zielzelle aus (Bukowski et al., 1985; Janeway et al., 2009; Kayser et al., 2005). Ein weiterer Bestandteil der angeborenen Immunabwehr ist das Komplementsystem, welches aus Plasmaproteinen und proteolyischen Enzymen besteht. Durch Lyse, Opsonierung und Einleitung von lokalen Entzündungsreaktionen können Bakterien und Viren inaktiviert und zerstört werden (Frank und Fries 1991; Rambach et al., 2008; Ricklin et al., 2010; Tosi 2005). Diese Art der Immunantwort erfolgt zeitnah und liegt somit vor den spezifischen Immunreaktionen (Fearon und Locksley 1996). Sollten die Erreger diese Front des angeborenen Immunsystems überwinden oder umgehen, ist eine spezifische, erworbene (adaptive) Immunantwort notwendig, wobei im Verlauf antigenspezifische Effektorzellen und Gedächtniszellen entstehen. Durch diese Eigenschaft des Immunsystems können Erreger, mit denen sich der Körper bereits zu einem früheren Zeitpunkt auseinandergesetzt hat, schnell erkannt und anschließend eliminiert werden. Dies steigert die Effektivität der immunologischen Prozesse und verhindert unnötige Begleitschäden. Das adaptive Immunsystem wird durch B- und T-Lymphozyten repräsentiert. 9 Einleitung Die B-Lymphozyten entwickeln sich aus den hämatopoetischen Stammzellen des Knochenmarks (B = bone marrow). Sie differenzieren sich zu Gedächtniszellen oder zu antikörpersezernierenden Plasmazellen. Sie sind in der Lage, mit oder ohne Kontakt Antikörper (Ig, zu T-Zellen (T-Helferzellen, Immunglobuline) zu bilden, Th-Zellen), wodurch antigenspezifische sie zur humoralen Immunantwort gezählt werden (Janeway et al., 2009; Neumann 2008). Neuere Studien zeigen jedoch auch einen negativen Effekt der B-Zellen auf das Immunsystem. Regulatorische B-Zellen sollen im Tiermodell das Immunsystem sogar unterdrücken und an der Entstehung einiger Krankheiten wie chronischer Colitis, Arthritis und autoimmuner Encephalitis beteiligt sein (Bouaziz et al., 2008; Matsushita et al., 2008; Mauri und Bosma 2012; Mauri und Ehrenstein 2008; Mizoguchi und Bhan 2006; Noh und Lee 2011; Yanaba 2009; Yang et al., 2009). T-Lymphozyten werden in der fetalen Leber und dem Knochenmark gebildet und gelangen über das Blutsystem in den Thymus (T-Lymphozyten), wo sie heranwachsen und ausreifen (Graf 2008). Durch Rekombination von Gensegmenten werden spezifische Rezeptoren für bestimmte Antigene gebildet. Je nach Differenzierungsgrad und Aktivierungszustand exprimieren die Lymphozyten auf ihrer Oberfläche zelluläre Glykoproteine, die „clusters of differentiation“ (CD) genannt werden (Geenen et al., 2001). So differenzieren TLymphozyten zu verschiedenen Populationen wie T-Helferzellen (Th-Zellen), die den Oberflächenmarker CD4 tragen, oder zu den CD8-positiven (CD8+) cytotoxischen T-Lymphozyten (CTL). Nach Heranreifung werden diese Zellen in das sekundäre lymphatische Gewebe ausgeschwemmt, wo sie auf ihre jeweiligen Antigene treffen sollen. Diese Antigene müssen den T-Zellen durch BLymphozyten, dendritische Zellen oder Makrophagen, die als antigenpräsentierende Zellen (APZ) bezeichnet werden, präsentiert werden. In den APZ lysierte Peptidfragmente der Antigene werden an bestimmte Oberflächenproteine, wie z. B. dem Haupthistokompatibilitätskomplex II (MHC = Major Histocompatibility Complex) gekoppelt, von den T-Zell Rezeptoren (TCR = tcell receptor) erkannt und gebunden (Fraser et al., 1998). Es werden zwei Klassen von Proteinen des MHC unterschieden. MHC I kommt auf der Oberfläche der meisten Zellen des Organismus vor und wird von den CD8- Rezeptoren der CTL’s gebunden. Die an MHC I gebundenen Antigene werden durch die CTL’s erkannt und vernichtet (Berke 1994; Kirchner et 10 Einleitung al., 1998). Darüber hinaus werden durch Aktivierung der CTL’s verschiedene Zytokine wie z. B. Interferone (IFN) freigesetzt, die unter anderem die antigenpräsentierenden Eigenschaften der APZ verstärken und eine Inflammation einleiten (Kirchner et al., 1998). Der von B-Lymphozyten und Makrophagen exprimierte MHC II wird von CD4-positiven (CD4+) T-Lymphozyten gebunden (Long 1992), wodurch eine Aktivierung mit anschließender Ausschüttung von Zytokinen resultiert. Anhand der Zytokinmuster werden die T-Helferzellen in Th1 und Th2 unterteilt (Delves und Roitt 2000a; Delves und Roitt 2000b; Janeway et al., 2009). Der Umstand, dass CD4+ T-Lymphozyten nur von MHC II präsentierte Antigene und CD8+ T-Lymphozyten ausschließlich an MHC I gebundene Antigene erkennen, wird als MHC-Restriktion der T-Lymphozyten bezeichnet (Stevanovic 2002; Zinkernagel und Doherty 1974a; Zinkernagel und Doherty 1974b). Eine weitere Lymphozytensubpopulation, die erstmalig 1995 von Sakaguchi beschrieben wurde, stellen die natürlichen regulatorischen T-Zellen (nTreg) dar (Sakaguchi 2005). Sie machen bis zu 10% der peripheren CD4-positiven TLymphozyten aus und sind in der Lage CD25 zu exprimieren (Sakaguchi 2005; Sakaguchi 2011; Stephens et al., 2001). Bei ihnen wurden ausgeprägte immunmodulierende Eigenschaften in vitro und in vivo nachgewiesen. Sie spielen eine wichtige Rolle für die Aufrechterhaltung der Selbsttolleranz (Murakami et al., 2002; Sakaguchi 2004; Shevach 2000) und werden mit unterschiedlichen Th1und Th2-bedingten Autoimmunerkrankungen in Verbindung gebracht (Bollinger et al., 2010; Corthay 2009; Tang und Bluestone 2008). Ferner sollen nTreg-Zellen in der Entstehung von Immunantworten gegen Tumore (Onizuka et al., 1999), Infektionen (Belkaid et al., 2002) und bei Reaktionen nach allogenen Organtransplantationen beteiligt sein (Cohen et al., 2002). Zusätzlich verhindern sie die Reifung der DCs und unterdrücken die Funktion der natürlichen Killerzellen (Cederbom et al., 2000; Miyara und Sakaguchi 2007). Sie inhibieren die Proliferation, Aktivierung und Zytokinproduktion von CD4+- sowie CD8+-Zellen (Sakaguchi 2004; Thornton und Shevach 1998) und von B-Lymphozyten (Miyara und Sakaguchi 2007). 11 Einleitung 1.1.2 Th1/Th2-Immunbalance An der Steuerung des Immunsystems sind unter anderem Zytokine beteiligt. Hierbei handelt es sich um Mediatoren, kurzlebige Botenstoffe, die für die Kommunikation der Zellen untereinander unerlässlich sind. Zu ihnen gehören Tumornekrosefaktoren (TNF), Interleukine (IL), Interferone (INF), Wachstumsfaktoren und koloniestimulierende Faktoren (CSF). Diese werden in unterschiedlichen Konzentrationen vorwiegend von Leukozyten exprimiert und bewirken unterschiedliche Reaktionen des Organismus (Ibelgaufts 2012; Janeway et al., 2009). Je nach ihrer Funktion kann man sie in proinflammatorische Zytokine, wie z. B. IL-1, IL-6, TNF-α (Cavaillon 1994; Kirchner et al., 1998) und antiinflammatorische Zytokine, wie z. B. IL-4, IL-5, IL-10 einteilen (Fiorentino et al., 1991a; Fiorentino et al., 1991b). Mosmann und Mitarbeiter entdeckten, dass naive T-Lymphozyten nach ihrer Aktivierung zwei funktionell verschiedene Zytokinmuster exprimieren. Anhand der freigesetzten Zytokine klassifiziert man die T-Helferzellen als Th1- und Th2Zellen. Th1 Leitzytokine sind IL-2, IL-12, IFN-γ und TNF-α. Th2-Zellen sind vor allem durch Ausschüttung von IL-4, IL-5, IL-6, IL-10 und IL-13 charakterisiert (Han et al., 2012; Kasakura 1998; Mosmann 1991; Mosmann und Coffman 1989; Ramos et al., 2011; Romagnani 1991; Szczypka et al., 2012; Takato-Kaji et al., 2005). Th1-Zytokine wirken proinflammatorisch. Sie führen zur Freisetzung von Makrophagen, aktivieren zytotoxische T-Zellen und sind besonders wichtig bei der Abwehr intrazellulärer Erreger (Viren, Chlamydien, Mykobakterien, Pilze). Th2Zytokine wirken antiinflammatorisch. Sie hemmen die Makrophagenaktivität, aktivieren Mastzellen und eosinophile Granulozyten. Außerdem stimulieren sie antikörperproduzierende B-Lymphozyten, was die humorale Immunantwort unterstützt. Dieser Abwehrmechanismus reguliert unter anderem allergische Reaktionen des Organismus sowie die Bekämpfung extrazellulärer Parasiten (Kidd 2003; Mosmann 1991; Mosmann und Coffman 1989; Romagnani 1991). Für einen optimalen Ablauf der Immunantwort ist die Balance der Th1- und Th2Zytokine von entscheidender Bedeutung (Kidd 2003). Beim gesunden Menschen sollte sie ausgeglichen sein und sich nach Ablauf einer Immunreaktion erneut einstellen. Überschießende Th1-Antworten können zu einer übermäßigen 12 Einleitung Entzündungsreaktion bis hin zum septischen Schock führen. Ein Überwiegen der Th2-Zytokine begünstigt hingegen eine Immunschwäche. Eine physiologische Th2-Dominanz sorgt jedoch für eine erfolgreiche Schwangerschaft und lässt sich auch in den ersten Lebensjahren eines Kindes nachweisen (Herberth et al., 2010; Kasakura 1998). Zudem wird in der Literatur eine rhythmische Schwankung der Th1/Th2-Balance mit einem Maximum in der ersten Nachthälfte beschrieben (Besedovsky et al., 2012; Dimitrov et al., 2004b; Lange et al., 2006a). Der Zusammenhang zwischen einer Immundysbalance und diversen chronischen Leiden wird durch eine Vielzahl von Studien belegt. So sind sowohl Erkrankungen wie rheumatoide Arthritis, Diabetes mellitus Typ I, als auch Multiple Sklerose oder Morbus Crohn mit einer Dominanz der Th1-Zytokine assoziiert. Bei Überlegenheit der Th2-Zytokine werden vor allem allergische Reaktionen, Infektionen mit dem humanen Immundefizienz-Virus (HIV) und das erworbene Immundefektsyndrom (AIDS) beobachtet (Herberth et al., 2010; Janeway et al., 2009; Kayser et al., 2005; Neumann 2008; Romagnani 1991). Zur Regelung und Aufrechterhaltung dieser Immunbalance sind die Zytokine ausgefeilten Regelmechanismen durch das Endokrinum und durch das Nervensystem unterworfen, wobei der Psyche und dem Schlaf ebenfalls eine große Rolle zugeschrieben wird (Ader et al., 1995; Besedovsky et al., 2012; Dhabhar 2009; Forsythe 2012; Ricklin et al., 2010; Whitesman und Booth 2004). Dabei hemmen sich die Zytokine der Th1- und Th2-Reihe gegenseitig (McEwen et al., 1997; Romagnani 1991). IFN-γ inhibiert beispielsweise selektiv die Proliferation und Funktion von Th2-Zytokinen, hat aber keinen direkten Einfluss auf die Th1Zellen. Die Th2-Zytokine IL-4 und IL-10 wirken ihrerseits supprimierend auf die Produktion der Th1-Zytokine (Gajewski und Fitch 1988; Gajewski et al., 1989). Neuere Studien zeigen auf, dass natürliche regulatorische T-Zellen (nTreg) ebenfalls an der Regulierung der Zytokinbalance beteiligt sind. NTreg unterdrücken die Sekretion von IFN-γ, IL-2 und TNF-α, aber nicht die von IL-4, IL-6 und IL-10 (Bollinger et al., 2010; Stroopinsky et al., 2009). Um ihre suppressive Wirkung entfalten zu können, sind sie wiederum wesentlich auf die Präsenz und Unterstützung von IL-6 und IL-2 angewiesen (Doganci et al., 2005; Pandiyan et al., 2007; Pasare und Medzhitov 2003). Bollinger und Mitarbeiter demonstrierten, 13 Einleitung dass Hormone, insbesondere Cortisol und Prolaktin in Zusammenarbeit mit nTreg die zirkadiane Steuerung der T-Zell-Aktivität modulieren (Bollinger et al., 2010). 1.2 Endokrinum und das Immunsystem Die Wechselwirkung zwischen dem Endokrinum und dem Immunsystem ist ein wichtiges Forschungsgebiet der Medizin. Die genauen Zusammenhänge und Wirkmechanismen sind bis dato nicht vollständig geklärt. Bereits in den späten vierziger Jahren des letzten Jahrhunderts stand fest, dass einige Hormone starke Auswirkungen auf das Immunsystem haben. So wurden z. B. dem Steroidhormon Cortisol immunsuppressive Eigenschaften zugewiesen. Es wurde zur Behandlung der rheumatoiden Arthritis verwandt, wofür Hench und Kollegen im Jahr 1950 den Nobelpreis verliehen bekamen (Kendall 1950; Lloyd 2002; Schubert und Schussler 2009). Das Endokrinum zählt neben dem Nervensystem zum wichtigsten Steuerelement des Organismus. Als „Kommunikationsmittel“ dienen Botenstoffe, unter anderem Hormone, die von Zellen des Endokrinums sezerniert werden und am Zielort ihre Wirkung über bestimmte Rezeptoren entfalten. Die „oberste Kontrollinstanz“ ist der Hypothalamus, der über komplexe Regulationsmechanismen als Verbindungselement zwischen Endokrinum und Nervensystem fungiert. Seine Verbindung mit dem Hypophysenvorderlappen (HVL) und der Nebennierenrinde (NNR) nennt man Hypothalamus-Hypophysen-Nebennierenrinden-Achse (HHNAchse) (Abb. 1). Über die Ausschüttung des Corticoliberins (Corticotropinreleasing Hormone = CRH) im Hypothalamus wird die Freisetzung vom Adrenocorticotropin (adrenocorticotropes Hormons = ACTH) im Hypophysenvorderlappen (HVL) gefördert. ACTH regt die Nebennierenrinde unter anderem zur Produktion von Mineralokortikoiden (MK), Glukokortikoiden (GK) und Sexualhormonen an. Sie regulieren über negative Feedback-Mechanismen die Freisetzung von ACTH und CRH (Hermus et al., 1984; Suda et al., 1985; Suda et al., 1987; Suda et al., 1986) und wirken auch auf Vorgänge innerhalb des Immunsystems (Arlt und Hewison 2004; Bornstein et al., 2004; Forsythe 2012; Janeway et al., 2009; Kelley et al., 2007; Kidd 2003). 14 Einleitung Abbildung 1: Schematische Darstellung der HHN-Achse. Regelmechanismen zur Freisetzung von Cortisol. HT = Hypothalamus, CRH = Corticotropin Releasing Hormone, HVL = Hypophysenvorderlappen, NNR = Nebennierenrinde, IS = Immunsystem, Rote Pfeile = Inhibition, Grüne Pfeile = Induktion Zwei weitere Hormone, die einen wichtigen Einfluss auf das Immunsystem haben und deren Freisetzung aus dem HVL über sogenannte Releasing - Hormone des Hypothalamus stimuliert wird, sind das Prolaktin und das humane Wachstumshormon (GH = human growth hormone). Deren Produktion und Freisetzung wird ebenfalls durch negative Rückkopplungsmechanismen reguliert (Dimitrov et al., 2004a; Jara et al., 2006; Kelley 1988; Kelley et al., 2007). Nachfolgende Abbildung schematisiert die Regulationsmechanismen der GHSekretion. Abbildung 2: Schematische Darstellung der Regelmechanismen zur Freisetzung von GH. HT = Hypothalamus, GHIH = Growth Hormone Inhibiting Hormone, GHRH = Growth Hormone Releasing Hormone, GIT = Gastrointestinaltrakt, HVL = Hypophysenvorderlappen, GH = Growth Hormone, IGF = Insulin-Like Growth Factor, IS = Immunsystem, Rote Pfeile = Inhibition, Grüne Pfeile = Induktion Im Folgenden wird auf die zwei, in dieser Studie untersuchten Hormone Cortisol und GH eingegangen. 15 Einleitung 1.2.1 Das Glukokortikoid Cortisol Kortikoide sind eine Gruppe von Hormonen, die in den Zellen der Nebennierenrinde gebildet werden. Anhand ihrer Hauptwirkung werden sie in Glukokortikoide (GK) und Mineralokortikoide (MK) eingeteilt. GK fördern die Glukoneogenese der Leber durch Induktion verschiedener Enzyme und stellen dadurch die Energieversorgung des Organismus in Bedarfssituationen sicher, wohingegen MK an der Aufrechterhaltung des Salz- und Wasserhaushaltes beteiligt sind. Über das Renin-Aldosteron-Angiotensin-System (R-A-A-S) regulieren sie das Blutvolumen. Ihre Wirkung entfalten beide Steroide über zwei Rezeptortypen: die Glukokortikoid- (GR) und Mineralokortikoidrezeptoren (MR) (de Kloet et al., 1993; Funder 1986; Funder 2000; Joels und de Kloet 1994). Die wichtigsten Liganden für GR und MR sind Cortisol und Aldosteron, wobei Cortisol seine Wirkung über beide Rezeptoren vermitteln kann. Als Antagonisten dienen unter anderem der GR-Antagonist RU-486 und der MR-Antagonist Spironolacton (Castinetti et al., 2012; de Kloet et al., 1999), womit in dieser Studie die Rezeptorbindung komplett unterbunden wurde. Das Glukokortikoid Cortisol wird in der Zona fasciculata der Nebennierenrinde über verschiedene Zwischenschritte aus Cholesterin gebildet. Wie bereits beschrieben, unterliegt die Cortisolsekretion der hypothalamischen und hypophysären Regulation durch CRH und ACTH. Aber auch andere Hormone (Fehm et al., 1984), wie z. B. GH (Castro-Magana et al., 1983) und Katecholamine (Rivier und Vale 1983) können seine Freisetzung beeinflussen. Zudem ist Cortisol selbst in der Lage, seine Synthese auf jeder Ebene der Bildungskaskade zu regulieren (Abb. 1). Die Plasmahalbwertszeit des Cortisols beträgt ca. 90 - 120 Minuten. Nach Abbau durch die Leber werden die entstandenen Metabolite über die Niere ausgeschieden (Köhrle und Petrides 2007a). Die Cortisolfreisetzung unterliegt einem strengen zirkadianen Rhythmus mit einem Höchstwert in den frühen Morgenstunden und sinkenden Werten in den späten Abendstunden, beziehungsweise während des Schlafes (Ball et al., 2006; Dimitrov et al., 2009; 16 Einleitung Esteban et al., 1991; Gallagher et al., 1973; Migeon et al., 1956; Nichols und Tyler 1967; Perkoff et al., 1959; Rimmele et al., 2010). Der Nadir der Cortisolfreisetzung wird zwischen Mitternacht und zwei Uhr morgens erreicht. Die wichtigste Funktion des Cortisols besteht in der Einleitung der Glukoneogenese zur kurzfristigen Bereitstellung von Energie bei der Bewältigung von Stresssituationen (Köhrle und Petrides 2007a; Munck et al., 1984). Diese Situationen können unter anderem körperliche Anstrengung (Brandenberger und Follenius 1975; Brandenberger et al., 1982), psychischer Stress (Holl et al., 1984) oder geistige Anspannung sein (Kamezaki et al., 2012). Ein Cortisolanstieg kann sogar nach einer Mahlzeit erfolgen (Follenius et al., 1982). Um die Wirksamkeit zu verbessern und die Nebenwirkungen zu verringern, wurden im letzten Jahrhundert synthetische Cortisolderivate entwickelt. Einige wichtige Vertreter sind das kurzwirksame Prednisolon und die langwirksamen Kortikoide Bethametason und Dexamethason (DEX) (Kaiser und Kley 2002; Mutschler et al., 2008). 1.2.2 Einfluss von Cortisol auf das Immunsystem und die Th1/Th2-Balance Neben seiner katabolen Hauptwirkung hat Cortisol auch immunmodulierende Eigenschaften, die bereits in den vierziger Jahren des letzten Jahrhunderts zur Behandlung immunologischer Erkrankungen, wie z. B. der rheumatoiden Arthritis ausgenutzt wurden (Hench et al., 1949; Kendall 1950). Die immunsuppressive Wirkung des Cortisols wurde in den letzten Jahren gründlich untersucht (Besedovsky et al., 2012; Pinto et al., 2006; Skjolaas et al., 2002; Yeager et al., 2008). Cortisol hemmt unter anderem die Entwicklung von Monozyten sowie die Proliferation und Expression von Rezeptoren auf Makrophagen. Durch GK wird die Anzahl der zirkulierenden Lymphozyten verändert, die Lymphozytenakkumulation, besonders im entzündeten Gewebe gehemmt und eine Lymphozyten-Apoptose induziert (Dimitrov et al., 2009; Dobbs et al., 1996; McEwen et al., 1997). Studien zeigen, dass durch erhöhte GK-Konzentrationen die Produktion der Th1-Zytokine IL-2, INF-γ und TNF-α gehemmt wird. Zudem kommt es zu einem leichten Sekretionsanstieg der Th2-Zytokine IL-10 und IL-4 (Agarwal und Marshall Jr. 17 Einleitung 2001; Gleeson und Bishop 2005; Visser et al., 1998; Yeager et al., 2008). Der endgültige Umschwung des Zytokinmusters zugunsten von Th2 soll eher durch die Inhibition der Th1-Zytokine, als durch Aktivierung der Th2-Zytokine zustande kommen. Analog dazu führt eine erniedrigte GK-Konzentration, die z. B. durch Blockade der GK-Wirkung hervorgerufen wurde, zur stärkeren Entwicklung Th1spezifischer Zytokine (Agarwal und Marshall Jr. 2001; Bollinger et al., 2010; Brinkmann und Kristofic 1995; Daynes und Araneo 1989; Dhabhar 2009; Franchimont et al., 2000; Franchimont et al., 1998; Pinto et al., 2006; Schreiber 2011). 1.2.3 Das humane Wachstumshormon (GH) GH ist ein Peptidhormon und wird im Hypophysenvorderlappen gebildet. Seine Sekretion wird unter anderem vom Hypothalamus über Growth Hormone Releasing Hormone (GHRH) und Somatostatin gesteuert. Ein weiterer Stimulus für seine Freisetzung ist das Hormon Ghrelin, welches größtenteils von der Magenmukosa produziert und ins Blut sezerniert wird (Köhrle und Petrides 2007b). GH wird pulsativ abgegeben und unterliegt einem ausgeprägten zirkadianen Rhythmus, wobei seine höchste Konzentration im Tiefschlaf gemessen werden kann. Daneben ist die Sekretion geschlechts- und altersabhängig (Antonijevic et al., 1999; Baeza et al., 2008; Veldhuis et al., 1997). Sie nimmt mit zunehmendem Lebensalter ab. GH löst in der Leber, aber auch in der Muskulatur und im Fettgewebe die Freisetzung von Insulin-ähnlichen Wachstumsfaktoren (IGF = Insuline like growth factors) aus, die zusammen mit Somatostatin die wichtigsten Inhibitoren der GHRH- und GH-Sekretion darstellen (Krueger und Obal 1993; Nyberg 2000) (Abb. 2). Die Hauptwirkung des GH wird durch IGF vermittelt und besteht in der Regulierung des Wachstums. Aus einem Hyposomatotropismus im Kindesalter kann ein Zwergwuchs resultieren. Eine Überproduktion dagegen führt entweder zu Gigantismus oder im Erwachsenenalter zu Akromegalie (de Boer et al., 1996; Köhrle und Petrides 2007b; Schiebler 2005). 18 Einleitung 1.2.4 Einfluss von GH auf das Immunsystem und die Th1/Th2-Balance Wie auch Cortisol besitzt GH immunmodulatorische Eigenschaften. Ihm werden Interaktionen mit T- und B-Zellen zugeschrieben. So soll GH immunstimulierende Fähigkeiten wie Förderung des Thymuswachstums oder Antikörperbildung besitzen. Ebenso sollen Makrophagen aktiviert, die Sekretion von Antikörpern durch B-Zellen stimuliert und über direkte Wirkung auf Monozyten die Freisetzung von proinflammatorischen Zytokinen (IL-12, IFN-γ und TNF-α) gefördert werden (Chappel 1999; Kelley et al., 2007; Postel-Vinay et al., 1997; Tripathi und Sodhi 2009). Ferner soll GH die Aktivität der CTL und NK sowie die Expression von Rezeptoren auf immunkompetente Zellen begünstigen (Auernhammer und Strasburger 1995; Berczi 1994; Chappel 1999; Davila et al., 1987). Mehrere Studien belegen, dass durch GH die Anzahl der Th1-Zytokine gesteigert und die Th2-Aktivität sogar erniedrigt wird (Barbano et al., 2001; Liu et al., 2008; Mustafa et al., 1997; Takagi et al., 1998), wodurch die Th1/Th2-Balance zugunsten der proinflammatorischen Th1-Antwort verschoben wird. Eine neue Studie von Borrione und Mitarbeitern beschreibt jedoch einen unerwarteten Shift zur Th2-Dominanz nach Gabe hoher Dosen des Wachstumshormons bei gesunden Sportlern (Borrione et al., 2012). 1.3 Schlaf, Endokrinum und das Immunsystem Ein weiterer Schwerpunkt der heutigen immunologischen Forschung ist die Wechselwirkung zwischen Immunsystem, Endokrinum und Schlaf. In vorangegangenen Studien wurde der regulatorische Einfluss des Schlafes auf immunologische Prozesse belegt. Schlaf gilt nunmehr als essenziell zur Aufrechterhaltung aller homöostatischer Prozesse, indem er auf den Stoffwechsel, das Endokrinum und das Immunsystem wirkt (Besedovsky et al., 2012; Horne 1988). In der Polysomnographie werden 5 Schlafstadien unterschieden. Das erste Stadium (S1) schließt sich direkt an die Einschlafphase an. Darauf folgt eine weitere Phase des leichten Schlafes (S2), indem der Körper noch ohne Weiteres 19 Einleitung erweckbar ist. Als Tiefschlaf oder slow wave sleep (SWS) werden die Stadien 3 und 4 bezeichnet, die sich durch langsamere Wellen mit hoher Amplitude auszeichnen. Diese Tiefschlafphasen findet man vor allem in der ersten Nachthälfte. Im zweiten Abschnitt der Nacht werden sie durch vermehrt auftretende und länger andauernde rapid-eye-movement Phasen (REM) abgelöst. Die Übergänge dieser Schlafphasen sind fließend und werden mehrmals pro Nacht durchlaufen (Rechtschaffen und Kahles 1968). Die im Rahmen dieser Studie untersuchten Hormone Cortisol und GH unterliegen einem schlafassoziierten Sekretionsmuster. Somit beeinflusst Schlaf deren Plasmakonzentration und deren Wirkung auf die Zytokinproduktion der T-Zellen. SWS-reiche Schlafphasen der ersten Nachthälfte inhibieren die Cortisolfreisetzung, was in einem Nadir zwischen 24:00 und 2:00 Uhr resultiert. Mit Abnahme der Tiefschlafphasen in der zweiten Nachthälfte steigen die CortisolPlasmaspiegel kontinuierlich an und erreichen ihr Maximum in den frühen Morgenstunden (Born und Fehm 1998; Gallagher et al., 1973; Weitzman 1976a; Weitzman 1976b). Auch bei GH wurde eine schlafassoziierte Abhängigkeit des Sekretionsmusters beobachtet. Im Gegensatz zu Cortisol wurde jedoch der Sekretionsgipfel in den ersten SWS-Phasen nach dem Einschlafen beschrieben. Im Verlauf der folgenden 24 Stunden lassen sich nur noch länger andauernde Phasen einer verminderten Freisetzung verzeichnen (Davidson et al., 1991; Lange et al., 2006a; Van Cauter und Copinschi 2000; Weitzman et al., 1981). Die Zusammenhänge zwischen Schlafstadien, Hormonfreisetzung und Zytokinbalance sind in der folgenden Abbildung dargestellt (Abb. 3). 20 Einleitung Abbildung 3: Schematische Darstellung eines Hypnogramms mit Cortisol- und GHPlasmakonzentrationen sowie dazugehörigem Zytokinprofil: W = Wachphase, REM = Rapid Eye Movement, S1 bis S4 = Non REM Schlafphasen, S3 und S4 = SWS = Tiefschlafphasen, GH = Wachstumshormon, Blaue Linie = Konzentrationverlauf GH, Rote Linie = Konzentrationsverlauf Cortisol, Grüne Fläche = Th1-Zytokinprofil, Gelbe Fläche = Th2-Zytokinprofil, GG = Th1/Th2Gleichgewicht (Balance) Wie in der obigen Skizze verdeutlicht, unterliegt die Zytokinsekretion diurnalen Schwankungen. Unter erniedrigtem Cortisolspiegel und hohen Konzentrationen von GH, wie sie in der ersten Nachthälfte (SWS-Phasen) bestehen, werden proinflammatorische Zytokine freigesetzt. Es kommt zur Sekretion von IL-1, IL-12, TNF-α und IFN-γ. Das Verhältnis der IFN-γ zu IL-4 produzierenden T-Zellen verschiebt sich zugunsten des Th1-Zytokins IFN-γ (Besedovsky et al., 2012; Dimitrov et al., 2004b; Esquifino et al., 2004; Hattori 2009; Haus 2007; Kelley et al., 2007). In der zweiten Nachthälfte und den frühen Morgenstunden, die durch erniedrigte GH- und erhöhte Cortisolspiegel bei vermehrt auftretenden REMPhasen gekennzeichnet sind, ist ein Umschwung der Th1/Th2-Balance zugunsten von Th2 zu verzeichnen (Besedovsky et al., 2012; Dimitrov et al., 2004b). Diese Zusammenhänge Immunsystem verdeutlichen und Schlaf, die wobei Interaktionen der zwischen Hypothalamus Endokrinum, als zentrales Regulationsorgan anzusehen ist. Er moduliert über den Ncl. suprachiasmaticus die zirkadiane Rhythmik, über Releasing- und Inhibiting-Hormone das Endokrinum, über den Locus coeruleus und Raphe Kerne den Schlaf sowie über Ausschüttung bestimmter Neurotransmitter das vegetative Nervensystem (Abb. 4). 21 Einleitung Abbildung 4: Schematische Darstellung der Immunmodulation: Wechselwirkungen und Rückkopplungsmechanismen zwischen Hypothalamus als Vermittler und zentraler Instanz zwischen Nervensystem, Endokrinum, zirkadianem Schrittmacher und Immunsystem (Guillemin 2005). HT = Hypothalamus, HVL = Hypophysenvorderlappen, EP = Epiphyse, CRH = Corticotropin Releasing Hormone (Corticoliberin), ACTH = Adrenocorticotropes Hormon, GHRH = Growth Hormon Releasing Hormon, GHIH = Growth Hormon Inhibiting Hormon SWS = Slow Wave Sleep = Tiefschlaf, REM = Rapid Eye Movement, NNR = Nebennierenrinde, GH = Growth Hormon, Th1/Th2 = Zytokine der Th1- und Th2-Helferzellen, Rote Pfeile = Inhibition, Grüne Pfeile = Induktion, Schwarze Pfeile = gegenseitige Beeinflussung Dem Wachstumshormon schreibt man proinflammatorische Effekte zu, während Cortisol konzentrationsabhängig eher antiinflammatorische Reaktionen hervorruft (Elenkov 2004; Kelley et al., 2007). Weitere schlafassoziierte Hormone sind Melatonin und Prolaktin. Auch sie sollen synergistische Effekte auf die Regulierung einer Immunantwort haben. Mit Anstieg ihrer Sekretion während des 22 Einleitung Schlafes sollen sie eine proinflammatorische Reaktion begünstigen (Besedovsky et al., 2012; Dimitrov et al., 2004b). Zusammenfassend besitzen der Schlaf, ebenso wie die zirkadiane Rhythmik, einen starken Einfluss auf das Immunsystem. Ein geregelter Schlaf-WachRhythmus ist essenziell für eine suffiziente Immunabwehr. Insomnie führt zu einer herabgesetzten Immunantwort und früher oder später zur Beeinträchtigung der Gesundheit (Besedovsky et al., 2012; Cohen et al., 2009; Dhabhar 2009; Everson 1993; Everson 2005; McEwen 2006; Meerlo et al., 2008). 23 Einleitung 1.4 Fragestellung und Ziel der Arbeit Diese in vitro Studie beschäftigt sich mit den Auswirkungen physiologischer Konzentrationen des humanen Wachstumshormons (GH) und des Cortisols auf das Immunsystem, insbesondere auf die Balance der Th1/Th2-Zytokine. Als Parameter für die Funktion des Immunsystems dienten IFN-γ- und IL-4produzierende CD4+ T-Helferzellen, wobei IFN-γ als Leitzytokin für Th1 und IL-4 für Immunreaktionen der Th2-Zellreihe dienten. Unter standardisierten Bedingungen wurden T-Lymphozyten aus Vollblutproben von 15 gesunden jungen Männern auf ihre intrazelluläre Konzentration an den o.g. Zytokinen am Durchflusszytometer untersucht. Im Einzelnen sollten folgende Fragen beantwortet werden. 1. Welchen Einfluss besitzen die Hormone GH und Cortisol auf IFN-γproduzierende CD4-positive T-Helferzellen? 2. Welchen Einfluss besitzen die Hormone GH und Cortisol auf IL-4-produzierende CD4-positive T-Helferzellen? 3. Welche Auswirkungen haben veränderte GH- und Cortisolkonzentrationen auf die Th1/Th2-Balance? 24 Material und Methoden 2. Material und Methoden 2.1 Probanden An dieser Studie nahmen 15 physisch und psychisch gesunde männliche Probanden im Alter zwischen 20 und 35 Jahren teil. Voraussetzung für die Teilnahme war ein gesunder und regelmäßiger Schlaf-Wach-Rhythmus, der einen Nachtschlaf von sieben bis neun Stunden ermöglichte und Schichtarbeit in den der Untersuchung vorausgegangenen sechs Wochen ausschloss. Innerhalb der letzten drei Monate durften die Probanden unter keinerlei Medikamenteneinfluss gestanden haben. Weitere Ausschlusskriterien waren Nikotinabusus und allergische Erkrankungen. Akute und / oder chronische Krankheiten wurden mittels eines Anamnesebogens (s. Anhang), klinischer Untersuchung sowie einer Routine-Laborkontrolle ausgeschlossen. Die Laborparameter schlossen ein großes Blutbild (Leukozyten < 9/µl, C-reaktives Protein (CRP) < 6mg/l), Elektrolytund Plasmametabolitbestimmungen und Bestimmung des TSH- (Thyroid Stimulating Hormon) Wertes ein. Bei der vorliegenden Studie handelt es sich um ein DFG-gefördertes Projekt (FOR457). Das Studienprotokoll wurde im Vorfeld von der Ethikkommission der Universität zu Lübeck vorgelegt und genehmigt. Jeder Proband wurde ausführlich aufgeklärt. Von jedem Teilnehmer lag eine schriftliche Einverständniserklärung vor Beginn der Studie vor. 2.2 Reagenzien Das humane Wachstumshormon (GH), der GR-Antagonist RU-486 sowie der MRAntagonist Spironolacton wurden über Sigma-Aldrich (St. Louis, Minnesota, USA) erworben. Die monoklonalen GH-Antikörper (5801) wurden von Oy Medix Biochemica Antibody (Kaunianinen, Finnland) hergestellt. 25 Material und Methoden 2.3 Versuchsablauf Um eine Aussage über die immunologischen Effekte physiologischer Konzentrationen von GH und Cortisol treffen zu können, wurden Vollblutproben gesunder Männer analysiert. Nach in vitro-Stimulation mit Ionomycin und PMA, in An- und Abwesenheit von GH und dem Cortisol-Rezeptor-Antagonisten RU-486 und Spironolacton wurde mittels Durchflusszytometers die Zytokinproduktion CD4positiver T-Helferzellen untersucht. Als Kontrollgruppe diente eine nicht stimulierte Vollblutprobe eines jeden Probanden. 2.3.1 Zellstimulation und Zellaktivierung Jedem Probanden wurde durch eine Venenpunktion 0,5 ml venöses Vollblut in heparinisierte Monovetten (Firma Becton Dickinson, San Jose, Kalifornien, USA) entnommen. Die Blutentnahme erfolgte, gemäß Studienprotokoll, immer morgens gegen 8:00 Uhr durch die gleiche Person. Die Probanden waren zum Abnahmezeitpunkt nüchtern und befanden sich in sitzender Position. Das Blut wurde unverzüglich im Verhältnis 1:1 mit dem Kulturmedium RPMI-1640 unter Zusatz von Penicillin, Streptomycin, Glutamin, nicht essentiellen Aminosäuren und Natriumpyruvat in Falcon-Röhrchen (Becton Dickinson, San Jose, Kalifornien, USA) versetzt. Danach folgte eine sechsstündige Inkubation der Proben in einem Brutschrank bei 37°C und 5 %-igem CO2-Zusatz in An- bzw. Abwesenheit von 20 ng/ml GH, 8,3 µM RU-486, einer Kombination aus GH und RU-486, und 8,3 µM Spironolacton. Durch den Zusatz von RU-486 und Spironolacton wurde eine komplette Blockade der Cortisolwirkung erreicht, da Cortisol seine Wirkung bekanntermaßen über beide Rezeptortypen (GR und MR) entfalten kann. So wurde der physiologische Nadir der Cortisol-Plasmakonzentration, wie er in der ersten Nachthälfte vorliegt, simuliert. Eine weitere Kultur, ohne jeglichen Zusatz, diente als unbehandelte Kontrolle. Diese Vollblutprobe enthielt die physiologisch hohe Cortisolkonzentration, die üblicherweise in den frühen Morgenstunden vorherrscht. Für die letzten vier Stunden der insgesamt sechsstündigen Inkubation wurden die Proben mit jeweils 1µg/ml Ionomycin und 8 ng/ml PMA (Phorbol-12myristat-13-acetat) stimuliert. Eine Proteinausschüttung wurde durch Zugabe des 26 Material und Methoden Golgi-Inhibitors Brefeldin A in einer Konzentration von 10 µg/ml verhindert. Alle hier verwandten Reagenzien wurden von Sigma-Aldrich Co., St. Louis, Minnesota, USA hergestellt. 2.3.2 Durchflusszytometrische Analyse Nach abgeschlossener Stimulation der Zellen, wie unter 2.3.1 erklärt, erfolgte deren Markierung für die durchflusszytometrische Analyse. Dazu wurden direkt konjugierte Antikörper gegen die Oberflächenmarker CD3 mit dem Fluorochrom Allophycocyanin (APC) (CD3-APC) und die Oberflächenmarker CD8 mit dem Farbstoff Peridin-Chlorophyll (PerCP) (CD8-PerCP) verwandt (Becton Dickinson, San Jose, Kalifornien, USA). Die Markierung wurde unter Lichtschutz bei Raumtemperatur für 20 Minuten durchgeführt. Anschließend erfolgte die zehnminütige Lyse der Erythrozyten mit 2 ml einer FACS-Lyse-Lösung (BD FACS™ lysing solution, Becton Dickinson, San Jose, Kalifornien, USA). Nach Zentrifugation für 5 Minuten bei 500g wurde der Überstand entfernt. Um ein Eindringen der zytokin-spezifischen Antikörper in die Zellen zu ermöglichen, erfolgte eine zehnminütige Permeabilisierungslösung (BD Permeabilisierung FACS™ permeabilizing mittels FACS- solution, Becton Dickinson). Nach der Zellwaschung mit CellWash + 0,5% FCS (fetales Kälberserum, fetal calf serum) erfolgte die Inkubation mit den intrazellulären Antikörpern gegen die Zytokine IFN-γ mit dem Fluorochrom Fluoresceinisothiocyanat (FITC) (IFN-γ-FITC) und gegen die Zytokine IL-4 mit dem Farbstoff Phycoerythrin (PE) (IL4-PE). Um eine Differenzierung zwischen positiven und negativen Zellen zu ermöglichen, wurden Isotyp-Kontroll-Antikörper benutzt (Machura et al., 2007; Pala et al., 2000; Prussin 1997). Nach einer erneuten Wäsche wurden die Zellen mit einer phosphatgepufferten Salzlösung (Phosphate buffered saline, PBS) versetzt und umgehend im Durchflusszytometer (BD FACSCalibur flow cytometer, Becton Dickinson) anhand der Software BD CellQuest Pro (ebenfalls Becton Dickinson) analysiert. Bei der Durchführung der Durchflusszytometrie wurde strikt nach der Anleitung des Geräteherstellers vorgegangen. Bei jeder Analyse wurden mindestens 10.000 27 Material und Methoden Zellen innerhalb der CD3+-Region, die den T-Lymphozyten entspricht, gezählt. Die Schwelle für Zytokin-positive Zellen wurde anhand von Isotyp-KontrollAntikörpern ermittelt. Um eine Kontamination durch andere Zellpopulationen auszuschließen, wurde die Vorwärts- und Seitwärts-Streuung für Lymphozyten zusammen mit den CD3-positiven Zellen (CD3+) entsprechend eingestellt. Die für diese Untersuchung relevanten IFN-γ- und IL-4-positiven Zellen wurden als prozentualer Anteil der CD3+CD8- T-Helferzellen ausgedrückt. Da der sonst übliche Marker CD4 durch Stimulation durch PMA herunterreguliert werden kann (Petersen et al., 1992; Suzuki et al., 1991), wurde im Rahmen dieser Studie der Marker CD8 zur Bestimmung der T-Helferzellen ausgewählt. 2.3.3 Hormonbestimmung Für die Hormonbestimmung wurde das durch eine morgendliche Venenpunktion gewonnene Blut in Serum-Monovetten (Sarstedt, Nümbrecht, Deutschland) gefüllt. Nach sofortiger Zentrifugation wurde der Überstand gewonnen und bis zur Hormonbestimmung bei -20 °C aufbewahrt. Die Messung des humanen Wachstumhormons (GH) erfolgte durch einen Radioimmunoassay (RIA, Diagnostic Products Corporation, Los Angeles, Kalifornien). Die Sensitivität der Untersuchung betrug 0,9 ng/ml. Der Intra- und Interassay-Variationskoeffizient (CV) lag unter 5,9 bzw. 8,3%. Für die Cortisol-Bestimmung wurde ein enzymgekoppelter Immunadsorptionstest (EIA) der Firma Diagnostic Systems Laboratories, Sinsheim, Deutschland mit der Sensitivität 0,1 µg/dl herangezogen. Der Intra- und Interassay-Variationskoeffizient (CV) lag unter 5,0 bzw. 12%. 28 Material und Methoden 2.3.4 Statistische Auswertung der Ergebnisse Folgende Parameter wurden statistisch ausgewertet: - Anzahl der IFN-γ-produzierenden CD4+ T-Helferzellen und IL-4- produzierenden CD4+ T-Helferzellen als Leitzytokine der Th1- bzw. Th2Immunantwort - absolute Änderung IFN-γ-produzierenden CD4+ T-Helferzellen und IL-4produzierenden CD4+ T-Helferzellen nach Zusatz von GH, RU-486, GH und RU-486 sowie Spironolacton. Um starke interindividuelle Schwankungen der Absolutwerte auszugleichen und eine genauere Aussage über die Konzentrationsänderung der Zytokine treffen zu können, wurden diese Absolutwerte zusätzlich ins Verhältnis zur unbehandelten Kontrollgruppe gesetzt und anschließend statistisch ausgewertet. Hierdurch wurden nicht die Absolutwerte, sondern die tatsächliche Änderung der Zytokinproduktion erfasst. Darüber hinaus wurde das Verhältnis der Th1- zu Th2-Helferzellen (Th1/Th2Balance) nach Zusatz der o.g. Substanzen verglichen zur Kontrollgruppe ausgewertet. Nach Prüfung der Normalverteilung mit Hilfe des parameterfreien KolmogoroffSmirnov-Testes erfolgte die Evaluation der Änderung der Th1- und Th2-Zellen nach Zugabe der o.g. Substrate im Vergleich zur unbehandelten Kontrollgruppe. Hierfür wurde der zweiseitige t-Test nach Student für abhängige Proben verwandt. Alle Daten wurden als Mittelwerte ± SEM (Standard Error of Mean) angegeben. Die Auswertung erfolgte mit der Software IBM SPSS Statistics 18 für Mac OS/X (SPSS Incorporated, an IBM Company, Chicago, Illinois USA). Die Unterschiede wurden als signifikant betrachtet, wenn die Irrtumswahrscheinlichkeit p < 0,05 war. 29 Ergebnisse 3. Ergebnisse Im Rahmen dieser Studie wurden Vollblutproben von 15 gesunden Männern im Alter von 20 bis 35 Jahren auf die Veränderungen von IL-4- bzw. IFN-γKonzentrationen nach in vitro-Stimulation untersucht. Als Kontrolle diente stimuliertes Blut ohne Zusatz von GH oder Rezeptorantagonisten. In diesen Proben lagen physiologische morgendliche Hormonkonzentrationen vor. IFN-γ repräsentierte die Zytokinproduktion des Th1-Profils, IL-4 wurde als Marker für die Th2-Zytokingruppe herangezogen. Die für diese Untersuchung relevanten IFN-γund IL-4-positiven Zellen wurden, wie unter Material und Methoden beschrieben, als prozentueller Anteil der CD3+CD8- T-Helferzellen ausgedrückt. 3.1 Plasma-Hormon-Konzentration innerhalb der Kontrollgruppe Der Schwellenwert für eine mögliche Bestimmung der GH-Konzentration im Blut liegt bei 0,9 ng/ml. Bei keinem der Probanden (n=15) konnte dieser Wert erreicht werden. Der Schwellenwert für die Cortisol-bestimmung (durch EIA) ist 0,1 µg/dl. Die Cortisol-Konzentration im Blut der Probanden schwankte zwischen 10,3 und 29,6 µg/dl (20,19 ± 1,53) (Mittelwert ± SEM) (Abb. 5) 30 Ergebnisse Abbildung 5: Schematische Darstellung der Cortisol - Plasmaspiegel der Probanden in µg/dl 3.2 Auswirkungen auf IFN-γ-produzierende CD4+ T-Helferzellen Die gemessenen Werte (Absolutwerte) produzierenden CD4+ T-Helferzellen der Kontrollgruppe, der IFN-γ- bewegten sich zwischen 11,4 und 35,4 (22,1 ± 1,8%). Nach Zugabe von humanem Wachstumshormon (growth hormone, GH) konnte ein Anstieg der Zellen auf 23,1 ± 1,7% gemessen werden. Den größten Einfluss auf die Produktion der IFN-γ-produzierenden CD4+ Zellen hatte die Zugabe des Spironolactons, wodurch sich die Zellzahl auf 25,6 ± 2,1% erhöhte (p=0,007) (Tab. 1; Abb. 6). Kontrolle RU-486 GH + RU-486 GH Spironolacton 22,1 ± 1,8 24,5 ± 2,1 24,4 ± 2,0 23,1 ± 1,7 25,6 ± 2,1 p-Wert 0,002 * 0,0007 * 0,04 * 0,007 * Tabelle 1: Absolutwerte (gemessene Werte) der IFN-γ-produzierenden CD4+ T-Helferzellen (in %) nach Zugabe von RU-486, GH+RU-486, GH und Spironolacton nach Inkubation mit Ionomycin und PMA (MW ± SEM), * = signifikant (p < 0,05) 31 Ergebnisse Abbildung 6: Schematische Darstellung der Absolutwerte (Mittelwerte ± SEM) der IFN-γproduzierenden CD4+ T-Helferzellen nach Zugabe von RU-486, GH+RU-486, GH und Spironolacton (Spiro) im Vergleich zur Kontrolle, * = signifikant (p < 0,05) Nach Zugabe von GH (p=0,04), von RU-486 (p=0,002), Spironolacton (p=0,007) und der Kombination aus GH und RU-486 (p=0,0007) war ein signifikanter Anstieg der IFN-γ-produzierenden CD4+ T-Helferzellen zu verzeichnen (Abb. 6; Tab. 1). Betrachtet man die relative Änderung der Zellzahlen im Verhältnis zur Kontrolle, so war hier ebenfalls eine signifikante Zunahme der IFN-γ-produzierenden CD4+ T-Helferzellen nach Zusatz von GH, RU-486, Spironolacton und der Kombination von GH und RU-486 erkennbar. Der größte Anstieg der IFN-γ-produzierenden CD4+ T-Lymphozyten wurde auch hier nach Zugabe von Spironolacton gemessen (Tab. 2; Abb. 7). 32 Ergebnisse RU-486 GH + RU-486 GH Spironolacton 11,3 ± 2,9 11,0 ± 2,6 5,5 ± 2,9 12,9 ± 3,8 p = 0,002 * p = 0,0004 * p = 0,048 * p = 0,005 * Tabelle 2: Relativ zur Kontrolle stattgefundene Änderung der IFN-γ-produzierenden CD4+ THelferzellen (in %) nach Zugabe von RU-486, GH+RU-486, GH und Spironolacton nach Inkubation mit Ionomycin und PMA (MW ± SEM) und die dazugehörigen p-Werte, * = signifikant (p < 0,05) Abbildung 7: Schematische Darstellung der Auswirkungen (Mittelwerte ± SEM) von RU-486, GH+ RU-486, GH sowie Spironolacton (Spiro) auf IFN-γ-produziernde CD4+ T-Helferzellen verglichen mit der Kontrolle, * = signifikant 3.3 Auswirkungen auf IL-4-produzierende CD4+ T-Helferzellen Die Ergebnisse der Kontrollgruppe bewegten sich zwischen 0,21 und 2,56 % (1,08 ± 0,15 %). In Tab. 3 und Abb. 8 sind die gemessenen Absolutwerte und die dazugehörigen p- Werte nach Zugabe der einzelnen Substanzen ersichtlich. Nach Zugabe der jeweiligen Substanzen kam es in dieser Gruppe zu keiner signifikanten Veränderung der Anzahl der IL-4-produzierenden CD4+ THelferzellen. 33 Ergebnisse Kontrolle RU-486 GH + RU-486 GH Spironolacton 1,08 ± 0,15 1,10 ± 0,16 0,99 ± 0,14 1,06 ± 0,17 1,05 ± 0,16 p - Wert 0,6 0,1 0,8 0,2 Tabelle 3: Absolutwerte der IL-4-produzierenden CD4+ T-Helferzellen (in %) nach Zugabe von RU-486, GH+RU-486, GH, und Spironolacton nach Inkubation mit Ionomycin und PMA (MW ± SEM) und die dazugehörigen p- Werte Abbildung 8: Schematische Darstellung der Absolutwerte (Mittelwerte ± SEM) der IL-4produzierenden CD4+ T-Helferzellen nach Zugabe von RU-486, GH+RU-486, GH und Spironolacton im Vergleich zur Kontrolle Betrachtet man die relativ zur Kontrollgruppe stattgefundene Änderung der Anzahl IL-4-produzierender CD4+ T-Helferzellen, sieht man ebenfalls keine signifikanten Unterschiede (Tab. 4; Abb. 9). RU-486 GH + RU-486 GH Spironolacton 4,8 ± 6,4 -6,9 ± 3,5 9,0 ± 13,0 -5,7 ± 4,9 p = 0,4 p = 0,06 p = 0,6 p = 0,2 Tabelle 4: Relativ zur Kontrollgruppe stattgefundene Änderung der IL-4 produzierenden CD4+ THelferzellen (in %) nach Zugabe von RU-486, GH+RU-486, GH und Spironolacton nach Inkubation mit Ionomycin und PMA (MW ± SEM) und die dazugehörigen p-Werte 34 Ergebnisse Abbildung 9: Schematische Darstellung der Auswirkungen (Mittelwerte ± SEM) von RU-486, GH+ RU-486, GH sowie Spironolacton (Spiro) auf IL-4-produzierende CD4+ T-Helferzellen verglichen mit der Kontrolle 3.4 Auswirkungen auf das IFN-γ-/ IL-4-Gleichgewicht (Th1/Th2-Balance) Da in vivo die Th1/Th2-Balance eine bessere Aussage über die Immunkompetenz des Organismus zulässt, als die isolierten Absolutwerte der Zytokine, wurde im Rahmen dieser Studie das Verhältnis der IFN-γ- zu IL-4-produzierenden CD4+ Tzellen bestimmt. Betrachtet man die Relation der IFN-γ- zu IL-4-produzierenden CD4+ Zellen (Th1/Th2-Balance), so hatten alle Substanzen einen Anstieg der IFNγ Produktion zur Folge, wobei durch Zugabe von GH+RU-486 und Spironolacton signifikante Werte erreicht wurden (p=0,0008 bzw. p= 0,002) (Abb. 10; Tab. 5). RU-486 GH + RU-486 GH Spironolacton 10,3 ± 5,9 21,6 ± 5,0 8,24 ± 8,7 23,1 ± 6,3 p = 0,1 p = 0,0008 * p = 0,3 p = 0,002 * Tabelle 5: Relativ zur Kontrollgruppe erfolgte Änderung des Verhältnisses der IFN-γ- zu IL-4produzierenden CD4+ T-Helferzellen (in %) nach Zugabe von RU-486, GH+RU-486, GH und Spironolacton nach Inkubation mit Ionomycin und PMA (MW ± SEM) und die dazugehörigen pWerte, * = signifikant (p < 0,05) 35 Ergebnisse Abbildung 10: Schematische Darstellung der relativen Änderung (Mittelwerte ± SEM) des Verhältnisses IFN-γ- zu IL-4-produzierenden CD4+ T-Helferzellen zur Kontrolle in % nach Zugabe von RU-486, GH+RU-486, GH und Spironolacton, * = signifikant (p < 0.05) 36 Diskussion 4. Diskussion Die Modulation einer Immunantwort stellt einen wichtigen klinischen Aspekt bei der Behandlung und Prävention unterschiedlichster Erkrankungen dar. Sie unterliegt komplexen Steuermechanismen, welche weiterer Erforschung bedürfen. Zytokine fungieren als wichtige Akteure immunologischer Reaktionen und sind daher Gegenstand klinischer Untersuchungen. Besondere Aufmerksamkeit wird in diesem Zusammenhang dem Th1/Th2-Verteilungsmuster, der so genannten Th1/Th2-Balance geschenkt. Inzwischen ist bekannt, dass Schlaf über weitreichende Einflüsse auf die Modulation des Zytokinprofils und somit auf die Art der Immunantwort verfügt. Hier werden bestimmte Hormonkonstellationen ausgenutzt, die sich begünstigend auf proinflammatorische Reaktionen auswirken (Besedovsky et al., 2012; Born et al., 1997; Dimitrov et al., 2004b; Lange et al., 2010). In diesem Zusammenhang gehören Cortisol und GH zu den immunologisch aktiven, schlafassoziierten Hormonen. In der vorliegenden Arbeit sollte die Auswirkung physiologischer Konzentrationen von Cortisol und GH auf das menschliche Immunsystem, insbesondere auf das Gleichgewicht der Th1/Th2-Zytokine analysiert werden. Dabei sollten Hormonzustände simuliert werden, die üblicherweise während der ersten Nachthälfte (SWS - Phasen) in vivo existieren. Mittels Durchflusszytometers wurden die Vollblutproben von gesunden männlichen Probanden auf Alterrationen bezüglich der Zytokinproduktion CD4+ T-Zellen untersucht. Als repräsentative Parameter der Th1- und Th2-Zellreihen dienten die Zytokine IFN-γ und IL-4. Die Wirkung des GH wurde direkt durch dessen Zugabe und die des Cortisols indirekt durch Blockade der Glukokortikoid- und Mineralokortikoidrezeptoren gemessen. IFN-γ wurde als repräsentatives Zytokin für das Th1-Profil gewählt. Es unterstützt als Hauptzytokin der proinflammatorischen, Th1-vermittelten Immunantwort die Proliferation von T-Helferzellen der unspezifischen zellulären Immunabwehr (Fearon und Locksley 1996; Gajewski und Fitch 1988; Gajewski et al., 1989; Maggi et al., 1992; Mosmann und Coffman 1989; Sad und Mosmann 1994). IFN-γ wurde in früheren Untersuchungen mehrfach als reproduzierbarer Parameter zur Evaluation der Th1-Funktionen herangezogen (Kidd 2003; Petrovsky und Harrison 37 Diskussion 1998; Skjolaas und Minton 2002; Szczypka et al., 2012). Analog dazu liegen Studien vor, die IL-4 als Leitzytokin für die Beurteilung einer Th2-Immunantwort postulieren (Agarwal und Marshall Jr. 2001; Holland et al., 2008; Romagnani 1991; Szczypka et al., 2012) und zudem bestätigen, dass dieses Zytokin für eine Th2-vermittelte Immunantwort unerlässlich ist (Chen et al., 2004; Liew 2002; Paul und Seder 1994; Swain et al., 1990). Diese Erkenntnisse führten zur entsprechenden Zytokinauswahl im Rahmen dieser Studie. Aber auch andere Zytokine können als repräsentative Parameter für eine Th1- oder Th2-gerichtete Immunantwort herangezogen werden. Dazu gehören unter anderem IL-2, IL-4, IL7, IL-10, TNF-α und IL-12 (Benedict et al., 2007; Besedovsky et al., 2012; Bollinger et al., 2009; Dimitrov et al., 2006; Dimitrov et al., 2007; Lange et al., 2010; Lange et al., 2006a; Lange et al., 2006b). In der vorliegenden Arbeit wurden ausschließlich gesunde, allergiefreie, männliche Nichtraucher einbezogen. Entsprechend den Protokollvorgaben wurden mittels eines Fragebogens (s. Anhang, Kapitel 7) und nach Bestimmung entsprechender Laborwerte, akute, chronische, psychische und medikamentös therapierte Erkrankungen ausgeschlossen. Probanden mit labordiagnostisch nachgewiesenen Infektionen (CRP über 6 mg/l und / oder erhöhter Leukozytenzahl > 9000/µl) wurden aus der Studie ausgeschlossen. Jede Infektion stellt eine relevante Stressbelastung für den Organismus dar (Dhabhar 2009; Dobbs et al., 1996; Kidd 2003) und führt unter anderem zu erhöhten Cortisolspiegeln und somit zu Veränderungen der Zytokinaktivität (Black 1994; Dhabhar 2009; Lucey et al., 1996; Pinto et al., 2006; Tosi 2005). Schlafstörungen oder regelmäßiger Konsum von Drogen und Alkohol haben gleichermaßen Auswirkungen auf die immunologische Situation des Probanden. Um verfälschten Ergebnissen entgegen zu wirken, wurden betroffene Probanden von der Teilnahme ausgeschlossen (Besedovsky et al., 2012; Born et al., 1997; Dimitrov et al., 2004b; Jerrells 1991; Lange et al., 2010; Redwine et al., 2003). Bei psychischen Leiden sind ebenso Alterationen in der Th1/Th2-Balance beschrieben. So wurden bei Untersuchungen schizophrener Patienten diskrepante Veränderungen der IFN-γ-Spiegel beobachtet (Schwarz et al., 2001). Auch bei depressiven Patienten fielen supprimierte IFN-γ-Spiegel bei parallel gesteigerter IL-4-Synthese auf (Myint et al., 38 Diskussion 2005; Song et al., 2009), so dass auch diese Patientengruppe aus der Studie ausgeschlossen wurde. Durch eine Begrenzung des Alters der Teilnehmer auf maximal 35 Jahre wurden altersbedingte Schwankungen der Immunsituation berücksichtigt. So wurde zum einen eine Vergleichbarkeit der Probanden untereinander sowie zu Ergebnissen anderer Studien ermöglicht. Diverse Autoren beschrieben eine Dysbalance des Immunprofils zugunsten von Th2 mit zunehmendem Alter (Castle et al., 1997; Rink et al., 1998; Schwab et al., 1992; Van Cauter et al., 2000). Aufgrund erhöhter hormoneller Schwankungen wurden Frauen aus der Studie ausgeschlossen (Kalu et al., 2008; Kruse et al., 2000; Piccinni und Romagnani 1996; Piccinni et al., 2000). Zudem stellten Autoimmunerkrankungen, z. B. der Schilddrüse, ein Ausschlusskriterium dar. Pathologisch veränderte Hormonwerte, z. B. TSH beeinflussen nachweislich die Immunkompetenz (Kelley et al., 2007; Kidd 2003; Klecha et al., 2008) und könnten dadurch zu nicht repräsentativen Ergebnissen führen. Die oben aufgeführten Kriterien bekräftigen die Wahl der Probandengruppe, um möglichst unverfälschte Voraussetzungen für diese Studie zu schaffen. Die im Blut der Probanden gemessenen Cortisolspiegel lagen innerhalb der üblichen Referenzbereiche. Die interindividuellen Schwankungen könnten durch geringe stressbedingte Konzentrationsänderungen bedingt sein. Beispielsweise könnte eine Trypanophobie zu einer vermehrten Sekretion des Stresshormons Cortisol führen (Kamezaki et al., 2012; Petrovsky und Harrison 1998; Schubert und Schussler 2009). Die GH-Konzentration in den morgendlichen Blutproben lag, wie erwartet, unterhalb der Detektionsgrenze (Dimitrov et al., 2004b; Lange et al., 2010; Lange et al., 2006a)(Kapitel 3.1). Ein Teil der immunologischen Forschung beschäftigte sich mit Zytokinen und ihren Auswirkungen auf das Immunsystem. Es gibt mehrere in Studien erprobte Methoden zur Untersuchung und Bestimmung von Zytokinen. Hierzu zählen unter anderem der Enzyme linked immuno-sorbant-assay (ELISA), Enzyme linked immuno-spot-assay (Elispot), Radio immuno-assay (RIA), Reverse-transcriptionpolymerase chain reaction (RT-PCR), die Westernblot-Technik und Verfahren zur Detektion einer messenger (m)RNA Expression (Arora 2002; Ford 2010; Pala et 39 Diskussion al., 2000; Skapenko und Schulze-Koops 2007). Im vorliegenden Studiendesign wurde die Untersuchung von stimulierten und aktivierten Vollblutproben am Durchflusszytometer gewählt. Der Nachweis reaktiv gebildeter Zytokine nach Antigenstimulation ist jedoch auch durch die Verwendung mononukleärer Zellen des peripheren Blutes (Peripheral Blood Mononuclear Cell, PBMC) möglich. In beiden Varianten werden die Zellen stimuliert, fixiert, permeabilisiert und die dann exprimierten Zytokine mittels monoklonaler fluoreszierender Antikörper markiert und anschließend am Durchflusszytometer dargestellt. Studien belegen, dass beide Verfahren vergleichbare Analysemethoden zur Zytokinmessung per Durchflusszytometer darstellen (De Groote et al., 1992; Jason und Larned 1997; McNerlan et al., 2002; Yaqoob et al., 1999), auch wenn in Untersuchungen von Waldrop und Mitarbeitern die Frequenz der Zytokinexpression bei Vollblutstimulationen im Vergleich zu PBMC höher war (Waldrop et al. 1997). In der vorliegenden Arbeit wurde die Vollblutanalyse verwandt, um ungewollte Zellaktivierungen, die eventuell durch konventionelle Separationsvorgänge verursacht werden könnten, zu umgehen. Eine Zell-Separation könnte eine Abtrennung der T-Zellen von ihrem umgebenden hormonellen Milieu bedeuten, welches jedoch gerade im Rahmen dieser Untersuchung von Interesse war (Schultz et al., 2002). Zudem wird durch Nutzung von Vollblutproben eine Reproduzierbarkeit sowie die Vergleichbarkeit mit bereits vorausgegangenen Studien unserer und anderer Arbeitsgruppen zu dieser Thematik ermöglicht (Dimitrov et al., 2009; Duramad et al., 2004; Lange et al., 2006a; Matalka und Ali 2005; Visser et al., 1998). Die im Rahmen dieser Arbeit verwandte Zytokinbestimmung mittels Durchflusszytometer ist eine anerkannte Methode zur Erfassung der Th1/Th2Immunaktivität und ermöglicht detaillierte Aussagen über das Zytokinmuster (Arora 2002; Brown und Wittwer 2000; Longobardi-Givani 1992; Pala et al., 2000). Die FACS setzt jedoch eine Stimulation der Zellen vor der Messung voraus, wodurch eine spontane Zytokinmessung nicht möglich ist. Ein weiterer Nachteil dieser Methode ist, dass zur intrazellulären Akkumulation der Zytokine Brefeldin A eingesetzt wird, welches zugleich verhindert, dass die stimulierten Zellen Rezeptoren exprimieren oder autokrine bzw. parakrine Mediatoren freisetzen. 40 Diskussion Andererseits erlaubt sie eine individuelle Analyse großer Zellpopulationen auf Einzelzellbasis (Jung et al., 1993; Pala et al., 2000) und mittels FACS-Analyse stellt sie die einzige Methode zur Identifizierung der gewünschten Zellreihen in heterogenen Suspensionen wie Vollblut dar (Kwak et al., 2000; Martin et al., 1992). Zudem ist die Durchflusszytometrie objektivierbar, d. h. vom Untersucher unabhängig und läßt die Vergleichbarkeit der Daten verschiedener Arbeitsgruppen zu (Brown und Wittwer 2000; Sack et al., 2000). Außerdem siedelt sich die Erfassungsgrenze für Zytokine unterhalb der für Serum-Testverfahren an (Lange et al., 2006a). Aus den oben aufgeführten Gründen wurde sich in dieser Studie für die FACS-Analyse als Mittel der Wahl zur Zytokinbestimmung und deren Quantifizierung entschieden. Die Interpretation durchflusszytometrisch ermittelter Daten ist von diversen Einflussfaktoren, wie Lagerung und Transport der Proben sowie Tageszeit und Körperlage zum Zeitpunkt der Blutabnahme, abhängig. Lange Transport- und / oder Lagerungszeiten können zu verfälschten Ergebnissen, gerade in der Bestimmung von Lymphozytenpopulationen, führen (Schultz et al., 2002; Tayebi et al., 1999). Für die korpuskulären Bestandteile im Vollblut ergeben sich niedrigere Werte in liegender Position der Probanden zum Zeitpunkt der Abnahme (Sack et al., 2000). Zudem sind für die in der vorliegenden Studie untersuchten Substanzen zirkadiane Rhythmen charakteristisch (Besedovsky et al., 2012; Born et al., 1997; Dimitrov et al., 2009; Lange et al., 2010; Levi et al., 1985; Levi et al., 1988). Um diesen Schwankungen entgegen zu wirken und vergleichbare Ergebnisse zu erzielen, wurde das Blut unter standardisierten Bedingungen entnommen und unverzüglich verarbeitet. Für die Stimulation der Zellen zur Zytokinproduktion wurden Ionomycin und PMA verwendet. Ionomycin und PMA sind unspezifische, unphysiologische Stimulantien des T-Zellrezeptor-Komplexes. Sie beeinflussen die intrazelluläre Bildung von Zytokinen wie IFN, IL-2 und IL-4 (Andersson et al., 1990; Rostaing et al., 1999; Yssel et al., 1992). In vivo hingegen benötigen T-Zellen ein spezifisches Antigen, welches nach Präsentation durch die APZ an den entsprechenden T-Zell-Rezeptor bindet und so zur Zytokinfreisetzung führt. Studien belegen aber, dass die Ergebnisse einer Zytokinstimulation in vivo durchaus mit denen einer Stimulation 41 Diskussion in vitro vergleichbar sind, auch wenn die letztgenannte APZ unabhängig durch unphysiologische Stimuli wie PMA / Ionomycin erfolgt (Jung et al., 1993; Schuerwegh et al., 2003). Somit stellt die in vitro-Stimulation von T-Zellen eine anerkannte, reproduzierbare und gut erforschte Methode dar (Lange et al., 2006a; Picker et al., 1995; Prussin 1997; Tayebi et al., 1999). Die PMA-Konzentration wurde mit 8 ng/ml relativ gering gehalten. Diese Dosierung entspricht jedoch der üblichen Standarddosierung zur Induktion einer Zytokinaktivität (McNerlan et al., 2002; Picker et al., 1995). Eine weitere Steigerung der PMA-Konzentration (25 ng/ml) führte in einer Pilotstudie und in einer parallelen Versuchsreihe unter analogen Bedingungen zu keiner signifikanten Änderung der Th1/Th2-Balance, was die Wahl der PMA-Dosierung bekräftigte (Dimitrov et al., 2004a; Lange et al., 2006a). Für die Blockade des Sekretionsapparates der Zellen und Freisetzung von Mediatoren wurde Brefeldin A (BFA) verwandt. Es führt zur intrazellulären Akkumulation der Zytokine und macht ihre intrazelluläre Messung mittels Durchflusszytometer erst möglich. Im Gegensatz zu Monensin, welches in einigen Studien zum gleichen Zweck herangezogen wurde, blockiert BFA den Proteintransport zwischen endoplasmatischem Retikulum und dem Golgi-Apparat. Nachweislich ist BFA der potentere Inhibitor der Exozytose bei geringerer Toxizität (Nylander und Kalies 1999; Schuerwegh et al., 2001). In der Literatur werden BFA-Konzentrationen von 0,1 bis 10 µg/ml beschrieben, wobei bereits bei einer Dosierung von 0,1 µg/ml eine suffiziente Inhibition des Golgiapparates nachgewiesen werden konnte (Lippincott-Schwartz et al., 1991). Um eine maximale Inhibition und eine Vergleichbarkeit mit der Literatur sowie vorangegangener und nachfolgender Studien unserer Arbeitsgruppe zu erreichen, wurde in diesem Versuch mit einer Endkonzentration von 10 µg/ml gearbeitet (Dimitrov et al., 2004a; Dimitrov et al., 2004b; Waldrop et al., 1997). In der vorliegenden Arbeit wurde die Auswirkung von Cortisol und GH auf die Produktion von CD4+ T-Zellen mit besonderem Augenmerk auf die Th1/Th2Zytokinbalance untersucht. Cortisol und andere Glukokortikoide sind für ihre immunsuppressiven und antiinflammatorischen Wirkungen bekannt und werden als Standardtherapeutika zur Behandlung immun- und entzündungsbedingter 42 Diskussion Erkrankungen eingesetzt (Belvisi und Hele 2003; Hench et al., 1950; Kendall 1950; Miner et al., 2005; Schreiber 2011). Heute mehren sich die Belege dafür, dass systemisch wirkende GK, darunter Cortisol, eher immunmodulierend agieren. Sie sollen selektiv die Th1-Antwort hemmen und eine humorale Immunreaktion fördern, wobei unter anderem vermutlich deren Dosierung eine erhebliche Rolle spielt. Während pharmakologisch eingesetzte, unphysiologische Konzentrationen die Immunreaktionen unterdrücken, sollen physiologische Dosen immunmodulierend wirken (Ashwell et al., 2000; Daynes und Araneo 1989; Elenkov 2004; Elenkov und Chrousos 1999; Franchimont et al., 2000; Sternberg 2001). Im Rahmen dieser Studie wurde die Wirkung des Cortisols auf die Zytokinproduktion und die Th1/Th2-Balance untersucht. Durch Blockade der GR durch RU-486 und MR durch Spironolacton wurden die Effekte des Cortisols unterbunden, wodurch ein Cortisol-Nadir, wie er während des Tiefschlafes besteht, simuliert wurde. Es ist bekannt, dass Cortisol seine Wirkung über Stimulation beider Rezeptoren entfaltet (De Kloet et al., 1998; Lange et al., 2006a; White et al., 1997). Die gewählte Dosis von 8,3 ηM RU-486 und 8,3 ηM Spironolacton zeigten in einer Pilotstudie eine komplette Blockade beider Rezeptoren. Wie man es aus den Ergebnissen entnehmen kann, fand durch die Blockade der MR durch Spironolacton eine signifikante Verschiebung der Th1/Th2-Balance in Richtung Th1 statt (Tab. 5; Abb. 10), was auf einen signifikanten Anstieg des Th1Zytokins IFN-γ durch IFN-γ-produzierende CD4+ T-Zellen zurückzuführen ist (Abb. 6, 7; Tab. 1, 2). Es erfolgte keine signifikante Änderung der IL-4-produzierenden CD4+ Th-Zellen durch die Blockade des MR (Tab. 3, 4; Abb. 8, 9). Im Gegensatz dazu wurde durch die Blockade des GR durch RU-486 weder die Th1/Th2Balance (Tab. 5; Abb. 10), noch die Anzahl IL-4-produzierender CD4+ Zellen signifikant verändert (Tab. 3, 4; Abb. 8, 9). Es kam jedoch zu einer signifikanten Zunahme der IFN-γ-produzierenden CD4+ Zellen (Abb. 6, 7; Tab.1, 2). Die Kombination von Wachstumshormon und dem Cortisolrezeptor-Antagonisten RU-486 führte zu einem signifikanten Shift in Richtung des Th1-Zytokinmusters (Tab. 5; Abb. 10), der auch in diesem Fall nicht auf einer Veränderung der IL-4produzierenden Th-Zellen (Tab. 3, 4; Abb. 8, 9), sondern durch den signifikanten 43 Diskussion Anstieg der IFN-γ-produzierenden CD4+ Zellen bedingt ist (Abb. 6, 7; Tab.1, 2). Zusammenfassend ist festzustellen, dass die Dysbalance zugunsten der Th1Zytokine unter vollständiger Inhibition der Cortisolwirkung auf einer signifikanten Zunahme der IFN-γ-produzierenden CD4+ Zellen beruht. Diese Ergebnisse werden von vielen in vitro-Studien indirekt bestätigt, in denen Cortisol einen Umschwung vom Th1- zum Th2-Profil vornehmlich durch Inhibition der Th1Zytokine bewirkt (Agarwal und Marshall Jr. 2001; Franchimont et al., 2000; Petrovsky und Harrison 1997; Petrovsky und Harrison 1998; Visser et al., 1998). Eine Zunahme des Th1-Profils unter abfallenden Cortisolwerten stellt eine Voraussetzung für inflammatorische Reaktionen dar, die im Rahmen einer suffizienten Abwehrreaktion notwendig sind. Diese Konstellation ist charakteristisch für die SWS-Phasen der ersten Nachthälfte und unterstreicht die Relevanz des Schlafes im Rahmen der Immunabwehr (Besedovsky et al., 2012; Born et al., 1997; Lange et al., 2010). Bei der Durchsicht der Literatur findet man kontroverse Aussagen über den Einfluss von Glukokortikoiden auf die Th2-Zytokine. Die Produktion dieser Zytokinreihe soll nach GK-Zusatz gesenkt, gesteigert oder unverändert bleiben. Nach Cortisoladministration fanden Lange und Mitarbeiter eine Reduktion der IL-4produzierenden CD4+ Zellen um 30% (Lange et al., 2006a), wobei andere Arbeitsgruppen eine dosisabhängige Reduktion von bis zu 90% beschrieben (Byron et al., 1992; Wu et al., 1991). Nach erneuter Stimulation einiger, bereits mit Glukokortikoiden anbehandelter Zellen fand eine signifikante Zunahme der IL-4Synthese statt (Agarwal und Marshall Jr. 1998; Agarwal und Marshall Jr. 2001; Brinkmann und Kristofic 1995; Daynes und Araneo 1989; Elenkov 2004; Ramirez 1998; Ramirez et al., 1996). Dies könnte entweder durch einen indirekten Effekt der GK, eine lange andauernde Stimulation oder die Anwesenheit von akzessorischen Zellen bedingt sein. Beispielsweise hemmen GK die Produktion von IL-12 durch Herabregulierung seiner Rezeptoren. IL-12, welches unter anderem von DC gebildet wird, induziert als potenter Inhibitor der IL-4-Zytokine und durch Steigerung der Synthese IFN-γ-produzierender CD4+ T-Zellen eine Th1-Dominanz. Eine Senkung des IL-12-Spiegels durch Cortisol führt demnach zu 44 Diskussion einem Anstieg der IL-4- und Reduktion der IFN-γ-produzierenden CD4+ Zellen, was in einer Verschiebung der Th1/Th2-Balance in Richtung Th2 resultiert (Besedovsky et al., 2012; Blotta et al., 1997; Dimitrov et al., 2007; Franchimont et al., 2000; Langenkamp et al., 2000; Moser und Murphy 2000). In der vorliegenden Studie konnte kein signifikanter Einfluss von GK auf IL-4 gezeigt werden, was durch die hier verwandten körpereigenen, physiologischen Konzentrationen für Cortisol bedingt sein könnte. Ein Großteil der Studien, in denen eine Änderung der IL-4-Produktion beschrieben wurde, benutzte zur Evaluierung der GK-Wirkung das potente, synthetische Glukokortikoid Dexamethason (DEX) in eher unphysiologischen Konzentrationen (Agarwal und Marshall Jr. 2001; Daynes und Araneo 1989; Franchimont et al., 2000; Franchimont et al., 1998; Ramirez 1998; Ramirez et al., 1996; Skjolaas et al., 2002; Visser et unphysiologischen al., 1998). Unter Konzentrationen Verwendung wurde synthetischer überwiegend eine GK in generelle Immunsuppression und Inhibition der Zytokinproduktion, sowohl der Th1- als auch der Th2-Zytokinreihe beobachtet (Braun et al., 1997; Kunicka et al., 1993; Snijdewint et al., 1995). Ein direkter Vergleich dieser Ergebnisse mit denen der vorliegenden Arbeit ist aufgrund der unterschiedlichen Potenz und Pharmakokinetik der verwandten GK nicht möglich. Beispielsweise liegt ein Großteil der körpereigenen GK in gebundener Form vor und nur ein Bruchteil des frei im Blut zirkulierenden Anteils ist biologisch aktiv (Tyrrell et al., 1986). Synthetische Steroide dagegen liegen vermehrt in ungebundener Form vor, was sich in ihrer höheren Wirkpotenz widerspiegelt (Fleshner et al., 2001; Pugeat et al., 1981; Skjolaas und Minton 2002; Spencer et al., 1990). Dies ist besonders bei in vivo-Studien zu bedenken. Ferner müssen hier bei der Interpretation und Vergleich der Ergebnisse synergistische oder antagonistische Effekte akzessorischer Zellen auf die Zytokinproduktion beachtet werden, die unter in vitro-Bedingungen ausgeschlossen werden können. Bei der Auswertung der Ergebnisse muss man sich mit der Wirkungsweise der Kortikosteroide und deren Rezeptoren auseinandersetzen. Kortikosteroide 45 Diskussion entfalten ihre Wirkungen über 2 Rezeptortypen, den Glukokortikoid- und Mineralokortikoid-Rezeptoren (Funder 1996; Funder 1997). Man ging davon aus, dass MR von Aldosteron, einem Mineralokortikoid aktiviert wird, worüber es seine Funktionen auf den Salz- und Wasserhaushalt des Körpers vermittelt. Analog dazu sollte das Glukokortikoid Cortisol seine Wirkung über GR entfalten (Frey et al., 2004). Heute ist bekannt, dass Cortisol seine Effekte über beide Rezeptoren vermittelt und sogar mit höherer Affinität an MR binden kann, als an GR (Funder 1997; Funder 2005; Mihailidou et al., 2009; Rupprecht et al., 1993). Abhängig von der An- und Abwesenheit des Enzyms 11β-Hydroxysteroiddehydrogenase (11βHSD) im betroffenen Gewebe bindet Cortisol vermehrt an MR als an GR (De Kloet et al., 1998; Edwards et al., 1996; White et al., 1997). Beide Rezeptoren sollen sogar als funktionelle Antagonisten fungieren (Funder 1993; Tanaka et al., 1997). Bei der Untersuchung der Th1/Th2-Balance und deren Beeinflussung durch GK wurde überwiegend auf DEX zurückgegriffen, welches selektiv an GR bindet. Bei der Interpretation einiger Studien, bei denen die Wirkung des Cortisols auf das Immunsystem eruiert wurde, ging man ebenfalls von seiner höheren Affinität zu GR aus (Dobbs et al., 1996). Die in dieser Studie durch Spironolacton blockierten MR bewirken einen signifikanten Umschwung des Gleichgewichts zugunsten der Th1-Zytokine. Der spezifische GR-Antagonist RU-486 induzierte einen ähnlichen Effekt, erreichte jedoch nur in Kombination mit GH-Signifikanz. Diese Beobachtungen könnten wie folgt interpretiert werden: 1. GR und MR arbeiten proinflammatorischer Zytokine bezüglich der synergistisch, cortisolinduzierten indem die Suppression Blockade beider Rezeptoren zu einer Steigerung des proinflammatorischen Zytokins IFN-γ und dadurch zu einer Verschiebung der Th1/Th2-Balance zugunsten von Th1 führte. Dieses Resultat widerspricht der Annahme, dass MR und GR als funktionelle Antagonisten fungieren (Funder 1993; Tanaka et al., 1997). Im Rahmen einer Parallelstudie wurde die Synergie beider Steroidrezeptoren durch Synthesesteigerung von TNF-α und IL-2 in CD4+ und CD8+ T-Zellen bei Blockade von GR und MR erneut bestätigt (Dimitrov et al., 2004a). 2. Cortisol entfaltet den größeren Teil seiner immunologischen Wirkung nicht, wie 46 Diskussion erwartet, über die GR, sondern über die MR. Auch Sauer und Mitarbeiter konnten durch MR-Blockade mittels Spironolacton in humanen Monozyten eine Umkehr der cortisolinduzierten Inhibition der IL-1-Sekretion, einem proinflammatorischen Zytokin, belegen (Sauer et al., 1996). Analog dazu konnten Vedder und Mitarbeiter einen Anstieg der TNF-α-Sekretion in menschlichen PBMC nach in vitro-Zusatz von Spironolacton feststellen (Vedder et al., 2003), was für die Wirkungsvermittlung der GK über MR spricht. In der oben genannten Parallelstudie konnte dieser TNF-α Anstieg über MR ebenfalls bestätigt werden (Dimitrov et al., 2004a). Zu diskutieren wäre an dieser Stelle außerdem auch der Einfluss des Mineralokortikoids Aldosteron auf das Immunsystem, denn dieses Hormon ist der eigentliche Ligand der MR. Aldosteron hat ebenfalls immunologisch modulierende Eigenschaften. Sie sollen unter anderem die Anzahl der zirkulierenden Lymphozyten, der NK und der Monozyten senken (Miller et al., 1994). Der periphere Plasmaspiegel von Cortisol liegt annähernd 1.000-fach über dem des Aldosterons (Charloux et al., 2001). Ferner liegt die Plasmaverweildauer des Aldosterons deutlich unter dem des Cortisols, da es kaum gebunden und bereits nach einer Leberpassage vollständig eliminiert wird. Die Plasmahalbwertszeit beträgt ca. 30 Minuten (Hubl et al., 1975). Des Weiteren findet man im Rahmen eines Hyperaldosteronismus einen Anstieg proinflammatorischer Zytokine wie z. B. TNF-α (Ahokas et al., 2003; Anker und Rauchhaus 1999; Carvajal et al., 2009). Trotz Blockade der MR durch Spironolacton im Rahmen der vorliegenden Arbeit kam es dennoch zu einem signifikanten Anstieg der proinflammatorischen Zytokine. Diese Feststellung und die oben aufgeführten Eigenschaften des Aldosterons sprechen gegen dieses Hormon als Auslöser der über MR-vermittelten Änderung der Zytokinbalance. Diese Argumente lassen die Schlussfolgerung zu, dass Cortisol seine Wirkung auf die Th1/Th2-Zytokinsynthese über MR ausübt. Ein weiteres Hormon, welches an der Immunmodulation beteiligt ist, ist GH. Ihm wird eine supportive Wirkung auf das Immunsystem zugeschrieben. Dieser Effekt wird entweder direkt von GH oder durch Insulin-like growth factor-1 (IGF1), dessen 47 Diskussion Bildung von GH stimuliert wird, ausgelöst (Auernhammer und Strasburger 1995; Clark 1997; Koo et al., 2001). Im Rahmen dieser Studie wurde der Einfluss von GH auf die Zytokinproduktion und die Th1/Th2-Balance untersucht. GH unterliegt einer starken, schlafgebundenen zirkadianen Rhythmik, mit einer maximalen Ausschüttung direkt nach dem Einschlafen und einem Nadir in der zweiten Nachthälfte und Morgenstunden (Born et al., 1997; Lange et al., 2010; Mallmann et al., 1989; Van Cauter und Copinschi 2000). Dies wurde durch die Messung der Hormonkonzentration im Rahmen dieser Studie bestätigt. Der GH-Spiegel im untersuchten Blut lag, wie erwartet, unterhalb der Bestimmungsgrenze (0,9 ng/ml). Um den Effekt einer GH Auswirkung auf das Immunsystem zu untersuchen, wurde den morgendlichen Blutproben GH in einer Konzentration von 20 ng/ml zugesetzt. Durch die Zufuhr des GH kam es zu einem signifikanten Anstieg des Th1-Zytokins IFN-γ. Eine signifikante Auswirkung auf die Zytokine der Th2-Reihe konnte jedoch nicht beobachtet werden. Ebenfalls konnte keine signifikante Änderung der Th1/Th2-Balance durch GH gemessen werden. Lediglich die Kombination von GH und RU-486 führte zu deren Verschiebung zugunsten von Th1 (Kapitel 3.1, 3.2, 3.3). Eine durch GH bedingte Steigerung der Th1-Zytokinproduktion wird durch die Ergebnisse mehrerer Arbeitsgruppen bestätigt (Lange et al., 2006a; Malarkey et al., 2002; Mellado et al., 1998; Mustafa et al., 1997; Sommese et al., 1996; Takagi et al., 1998). Eine signifikante Änderung der IL-4-produzierenden CD4+ Zellen konnte in dieser Studie nach GH-Applikation nicht festgestellt werden. Diese Resultate wurden ebenfalls durch mehrere Studien belegt (Galdiero et al., 1995; Galdiero et al., 1997). Im Widerspruch dazu lassen sich jedoch auch Berichte zu GH-bedingten signifikanten Änderungen der Th2-Zytokine, unter anderem von IL-4 finden (Liu et al., 2008). Die Arbeitsgruppe um Mellado beschrieb einen Anstieg der IFN-γ- aber einen Rückgang der IL-4-Produktion nach Zugabe von GH (Mellado et al., 1998). Takagi et al. haben einen signifikanten Rückgang von IL-4 durch GH an Mäusen, 48 Diskussion die einem Verbrennungstrauma ausgesetzt waren, festgestellt. Dieses führte zu einem Anstieg der Th2- und einem Abfall der Th1-Zytokinsynthese, repräsentiert durch IL-4 bzw. IFN-γ. Nach Zugabe von rekombinantem GH gelang eine Umkehr dieses Zytokinmusters erneut zugunsten von Th1, indem IFN-γ wieder anstieg und IL-4 sank (Takagi et al., 1998). Im Gegensatz zur vorliegenden Arbeit waren diese jedoch in vivo-Studien. Schlussfolgernd ist zu sagen, dass GH eine Th1-vermittelte Immunantwort induziert, welche in vivo, jedoch nicht in vitro, zu einer indirekten Suppression der Th2-Zytokine führen kann. Eine generelle Suppression der Th2-Immunantwort durch eine verstärkte IFN-γ-Aktivität wurde bereits mehrfach beschrieben (Gajewski und Fitch 1988; Gajewski et al., 1989). Ein weiterer Erklärungsversuch dieser Resultate könnte in der Verteilung der GHRezeptoren liegen. Die Expression der Rezeptoren für das Wachstumshormon auf T-Zellen ist gering, wobei fast alle (80-90%) B-Zellen und Makrophagen GHRezeptor-positiv sind (Bresson et al., 1999; Dardenne et al., 1998; Postel-Vinay et al., 1997). Dies lässt vermuten, dass der direkte Effekt von GH auf die T-Zellen geringer ausfallen muss, als seine indirekte Wirkung auf die T-Zellen, welche über die APZ vermittelt wird. Derartige indirekte Effekte des GH können in der vorliegenden Arbeit durch die APZ unabhängige Stimulation der Zellen mit PMA maskiert werden (Lange et al., 2006a). Im Gegensatz zu einem Abfall der Th2-Zytokine konnte Borrione einen signifikanten Anstieg selbiger durch GH feststellen. Da Borrione jedoch die Auswirkung unphysiologischer GH-Konzentrationen ab 200 ng/ml bis 600 ng/ml untersucht hatte, können diese Ergebnisse mit denen der vorliegenden Arbeit, in der physiologische Konzentrationen verwandt wurden, nicht verglichen werden (Borrione et al., 2012). Ein Anstieg der Th2-Zytokine durch GH wurde auch durch die Arbeitsgruppe um Ramos beobachtet. Hier wurde gezeigt, dass - GH die IL-10-Sekretion der CD4+ T-Zellen in gesunden Probanden stimuliert und - IL-10 die Sekretion der Th1-Zytokine verhindert, wodurch die Th1/Th2-Balance zugunsten von Th2 verschoben wurde (Ramos et al., 2011). 49 Diskussion Ein Vergleich ist ebenfalls nicht möglich, da im Rahmen der vorliegenden Arbeit die IL-10-Aktivität nicht untersucht wurde. Obwohl auch IL-10 zur Beantwortung der Fragestellung dieser Arbeit hätte dienen können, wurde sich aufgrund der Vergleichbarkeit mit vorangegangenen Studien unserer Arbeitsgruppe für IL-4 entschieden (Dimitrov et al., 2004a; Dimitrov et al., 2004b; Lange et al., 2006a). Zudem wurde eine mögliche immunmodulierende Wirkung von IGF-1, welches durch Stimulation von GH unter anderem von der Leber sezerniert wird, in dieser Studie nicht untersucht. In vivo hat dieser Wachstumsfaktor ebenfalls weitreichende Einflüsse auf die Immunkompetenz (Bernabei et al., 2003; Hattori 2009; Tu et al., 1999). Bei der Betrachtung der Auswirkungen der hier untersuchten Hormone auf das Immunsystem sollte auch die Rolle ihrer Releasing-Hormone diskutiert werden. Wie bereits erwähnt, wird die Freisetzung des GH durch GHRH reguliert (Abb. 1). GHRH selbst hat ebenfalls immunmodulierende Eigenschaften (Siejka et al., 2005a; Siejka et al., 2005b; Stepien et al., 2010; Valtorta et al., 1991), die zu Verschiebung der Th1/Th2-Balance führen können. Siejka und Mitarbeiter zeigten auf, dass GHRH unter in vitro-Bedingungen zu einem signifikanten Anstieg der IFN-γ-Sekretion in PBMC führt (Siejka et al., 2004). Die Freisetzung von Cortisol wird unter anderem über CRH reguliert. Auch dieses Hormon beeinflusst inflammatorische Reaktionen. Im Rahmen seiner supportiven Eigenschaften unterstützt es die Lymphozytenproliferation (Gay et al., 2008; Karalis et al., 1997; McGillis et al., 1989; Singh 1989). Über Aktivierung der HHNAchse durch die Zytokine TNF-α, IL-1 und IL-6 wirkt CRH über vermehrte Ausschüttung des Cortisols letztlich immunsuppressiv (Besedovsky et al., 1986; Sapolsky et al., 1987). Der weit verbreitete Spruch „Schlaf Dich gesund“ fand durch zunehmende Forschung auf den Gebieten der Neuroendokrinologie und Immunologie der letzten Jahrzehnte immer wieder Bestätigung. Schlaf dient also nicht nur der Erholung, sondern in gewissem Maße auch der zellulären Regeneration und 50 Diskussion unterstützt das Abwehrsystem. Dabei werden die vegetativen und hormonellen Gegebenheiten während des Schlafes ausgenutzt, um Pathogene suffizient zu bekämpfen. Klinische Beobachtungen verdeutlichen den engen Zusammenhang zwischen Schlaf und Immunsystem. Beispielsweise leiden Patienten mit Insomnie oder chronischen Schlafstörungen, genauso wie ältere Menschen, unter einer verstärkten Anfälligkeit für Infektionen mit protrahiertem Heilungsverlauf (Besedovsky et al., 2012; Zikorsky et al., 2001). Demgegenüber zeigten Lange und Mitarbeiter, dass gesunder Schlaf im Gegensatz zu Schlafentzug nach einer Hepatitis A-Impfung zu einer deutlich höheren Aktivität der antigenspezifischen THelferzellen führt. Diese Veränderung der Immunlage konnte ein Jahr nach erfolgter Impfung noch nachvollzogen werden (Lange et al., 2011). Um in diesem in vitro-Modell schlafassoziierte hormonelle Verhältnisse und ihre Einflüsse auf die Zytokinproduktion zu simulieren, erfolgte die Stimulierung und Aktivierung der morgendlichen Vollblutproben nach Studienprotokoll. Die für die SWS-Phasen charakteristischen Konzentrationen an GH und Cortisol wurden erreicht. Dennoch limitieren einige Faktoren die Interpretation der Ergebnisse. In vivo wird das Zytokinmilieu und die Aktivität dieser Mediatoren durch verschiedene Faktoren beeinflusst, welche jedoch unter experimentellen Bedingungen nicht reproduzierbar sind (Born et al., 1997; Dhabhar 2002; Dhabhar et al., 1994; von Andrian und Mackay 2000). Dennoch gehen die vorliegenden Ergebnisse mit denen anderer Arbeitsgruppen konform. Für die SWS-Phasen und für die erste Nachthälfte sind Sekretionssteigerungen der proinflammatorischen Zytokine charakteristisch. So beschrieben Petrovsky et al. ein Maximum der IFN-γ-Sekretion gegen 23 Uhr (Petrovsky und Harrison 1997). Dieses Ergebnis stimmt mit denen anderer Arbeitsgruppen überein, die für dieses Zytokin einen Höchstwert in der ersten Nachthälfte postulieren (Dimitrov et al., 2004b; Petrovsky und Harrison 1998; Petrovsky et al., 1998). Bisherige Forschungsergebnisse über die proinflammatorischen Zytokine IL-1, IL-6, TNF-α, IL-2 und IL-12 bestätigen das Vorliegen einer Th1-dominierten Immunantwort während des nächtlichen Schlafes (Born et al., 1997; Gudewill et al., 1992; Irwin et al., 2000; Palm et al., 1996; Petrovsky et al., 1998; Zabel et al., 1993). Dimitrov und Mitarbeiter beschrieben 51 Diskussion sogar einen Shift der Balance zugunsten von Th1, gemessen anhand der Zytokine IFN-γ und IL-4 (Dimitrov et al., 2004b). Dies bekräftigt sich erneut im Rahmen der vorliegenden Studie, indem unter einer schlafähnlichen Hormonkonstellation eine Th1-Dominanz, allein durch die isolierte Zunahme von IFN-γ-produzierenden CD4+ T-Helferzellen zu verzeichnen war. Die IL-4-Synthese wurde in diesem hormonellen Milieu nicht signifikant beeinflusst. Zum anderen zeigt es, dass Cortisol eine deutlich höhere immunmodulierende Potenz besitzt, als sein Gegenspieler GH. Dies wird deutlich, indem GH nur durch parallele Drosselung der Cortisolwirkung signifikante Effekte auf die Th1/Th2-Balance aufwies. Die Kernfragen der Studie konnten anhand der erzielten Ergebnisse wie folgt beantwortet werden: 1. Die Zugabe von GH in einer physiologischen Konzentration von 20 ng/ml zu den morgendlichen Blutproben der Probanden führte zu einem signifikanten Anstieg der IFN-γ-produzierenden CD4+ T-Helferzellen. Die Wirkung des Cortisols auf die IFN-γ-produzierenden CD4+ T-Helferzellen wurde durch Blockade der MR und GR untersucht. Dies führte zu einem signifikanten Anstieg der IFN-γproduzierenden CD4+ T-Helferzellen. Im Umkehrschluss führt Cortisol zu einem signifikanten Abfall der IFN-γ-produzierenden CD4+ T-Helferzellen. 2. Durch die Zugabe von GH in einer physiologischen Konzentration von 20 ng/ml zu den morgendlichen Blutproben der Probanden konnte kein signifikanter Effekt auf IL-4-produzierende CD4+ T-Helferzellen gezeigt werden. Ebenfalls konnte dem Cortisol durch Blockade von MR und GR kein signifikanter Einfluss auf die IL4-produzierenden CD4+ T-Helferzellen nachgewiesen werden. 3. Durch die Zugabe von GH in einer physiologischen Konzentration von 20 ng/ml zu den morgendlichen Blutproben der Probanden konnte kein signifikanter Effekt auf die Th1/Th2-Balance beobachtet werden. Lediglich durch die Blockade von MR durch Spironolacton wurde die Th1/Th2-Balance signifikant zugunsten von Th1 verschoben. Die Blockade von GR führte nur in Kombination mit GH zu einer 52 Diskussion signifikanten Änderung dieses Gleichgewichtes in Richtung Th1-Dominanz. Im Umkehrschluss führt Cortisol zu einem Umschwung der Th1/Th2-Balance zur Th2Dominanz, vermittelt durch MR. Unabhängig von den anfangs gestellten Fragen führte die Auswertung der vorliegenden Arbeit zu weiteren Kenntnissen, die sich wie folgt zusammenfassen lassen: 1. Die Verschiebung der Th1/Th2-Balance erfolgt durch die Auswirkung des Cortisols durch Änderungen der Th1-und nicht der Th2-Zytokinaktivität. 2. Cortisol entfaltet seine immunmodulierende Wirkung über MR und nicht über GR. 3. Erhöhte GH- sowie erniedrigte Cortisol-Konzentrationen bewirken eine vermehrte Synthese des Th1-Zytokins IFN-γ, die in einer Th1 dominierten Immunantwort resultiert. Im Rahmen dieser in vitro Studie ist es gelungen, schlafähnliche Hormonverhältnisse zu simulieren. Diese führten, wie erwartet, zu einer Verschiebung der Th1/Th2-Balance zugunsten der Th1-Zytokine. Dieses Zytokinprofil ist essenziell für die proinflammatorische Immunantwort im Rahmen einer Abwehrreaktion und spiegelt die Bedeutung des Schlafes bezüglich seiner immunmodulierenden Eigenschaften wider. Um die Aussagen der vorliegenden Arbeit zu verifizieren, sollten weiterführende Studien durchgeführt werden. Unter anderem wäre es sinnvoll, die Blutentnahme während der SWS Phasen durchzuführen, um zu eruieren, ob eine Reproduzierbarkeit der Ergebnisse möglich ist. Weiterhin könnten physiologische Stimuli der Zytokinsynthese, wie z. B. Staphylokokkus Enterotoxin B verwendet werden. 53 Zusammenfassung 5. Zusammenfassung Eine suffiziente Immunabwehr ist essenziell für das Überleben eines Organismus in einer Umwelt verschiedenster gesundheitlicher Herausforderungen. Ein ausgewogenes Verhältnis zwischen Th1-und Th2-Zytokinen, welches auf den immunologischen Reiz optimal abgestimmt sein sollte, ist hierbei von herausragender Bedeutung. Die Steuerung einer Immunantwort unterliegt komplexen Regulationsmechanismen, wobei das endokrine System eine wichtige Instanz darstellt. Hierbei übernehmen Cortisol und GH als Immunmodulatoren eine wichtige Rolle. Ziel der vorliegenden Arbeit war es, die Auswirkung physiologischer Konzentrationen des humanen Wachstumshormons (GH) und des Cortisols auf das Immunsystem, insbesondere auf die Balance der Th1/Th2-Zytokine unter in vitro-Bedingungen zu untersuchen. Als repräsentative Parameter dienten IFN-γbzw. IL-4-produzierende CD4+ T-Helferzellen. Die Wirkung des GH wurde direkt durch dessen Zugabe bestimmt, während die Messung der Cortisoleffekte indirekt über Blockade der Glukokortikoid- und Mineralokortikoidrezeptoren erfolgte. Vollblutproben von 15 gesunden jungen Männern wurden durch Ionomycin und PMA in An- und Abwesenheit folgender Substanzen stimuliert: GH, RU-486 (GRAntagonist), Kombination aus GH und RU-486 sowie Spironolacton (MRAntagonist). Die intrazelluläre Zytokinproduktion wurde mittels eines Durchflusszytometers gemessen. Unter Administration von GH wurde lediglich eine signifikante Steigerung der IFNγ-produzierenden CD4+ T-Helferzellen beobachtet. Die IL-4-produzierenden CD4+ T-Helferzellen sowie die Th1/Th2-Balance blieben unbeeinflusst. Die Kombination von GH und RU-486 führte zu einer signifikanten Zunahme der IFN-γ-produzierenden CD4+ T-Helferzellen, durch Ausbleiben einer signifikanten Änderung IL-4-produzierender CD4+ T-Helferzellen in einer signifikanten Th1Dominanz resultierte. Nach Blockade von MR und GR durch die Zugabe von Spironolacton bzw. RU-486 54 Zusammenfassung wurde eine signifikante Synthesesteigerung IFN-γ-produzierender CD4+ THelferzellen protokolliert. Jedoch führte keine der zugesetzten Substanzen zu einer signifikanten Änderung IL-4-produzierender CD4+ T-Zellen. In Bezug auf die Th1/Th2-Balance zog ausschließlich Spironolacton eine signifikante Verschiebung zugunsten der Th1-Dominanz nach sich. Zudem konnte gezeigt werden, dass Cortisol seine immunmodulatorische Wirkung hauptsächlich über MR und nicht über GR entfaltet. Die Dysbalance zugunsten des Th1-Zytokinprofils resultiert primär durch Zunahme des Th1-Zytokins IFN-γ- und nicht durch Veränderungen der IL-4-Synthese. Die oben genannten Erkenntnisse wurden durch Simulation schlafähnlicher Hormonkonstellationen gewonnen. Durch niedrige Cortisolspiegel und hohe GHKonzentrationen, die für die ersten Stunden des Schlafes charakteristisch sind, entstand ein proinflammatorisches Zytokinprofil durch eine gesteigerte IFN-γSynthese. Dies verdeutlicht die Notwendigkeit eines geregelten Schlaf-WachRhythmus für einen suffizienten und ungestörten Ablauf immunologischer Prozesse. Die Möglichkeit durch fundiertes Wissen auf dem Gebiet der Immunologie Abwehrreaktionen von extern beeinflussen oder gar steuern zu können, ist wichtiger Bestandteil der heutigen Forschung. Auch im Rahmen der Therapie von Immunerkrankungen zeigt sie neue Möglichkeiten auf. Bis dato sind jedoch nicht alle Zusammenhänge und Wirkmechanismen bekannt und bedürfen weiterer klinischer Erforschung. Besonderes Augenmerk sollte dabei den möglichen Therapieansätzen immunbedingter Erkrankungen gelten. 55 Literturverzeichnis 6. Literaturverzeichnis Ader R, Cohen N, Felten D: Psychoneuroimmunology: interactions between the nervous system and the immune system. Lancet 345(8942), 99-103 (1995) Aderem A, Underhill DM: Mechanisms of phagocytosis in macrophages. Annu Rev Immunol 17, 593-623 (1999) Agarwal SK, Marshall Jr. GD: Glucocorticoid-induced type 1/type 2 cytokine alterations in humans: a model for stress-related immune dysfunction. J Interferon Cytokine Res 18(12), 1059-68 (1998) Agarwal SK, Marshall Jr. GD: Dexamethasone promotes type 2 cytokine production primarily through inhibition of type 1 cytokines. J Interferon Cytokine Res 21(3), 14755 (2001) Ahokas RA, Warrington KJ, Gerling IC, Sun Y, Wodi LA, Herring PA, Lu L, Bhattacharya SK, Postlethwaite AE, Weber KT: Aldosteronism and peripheral blood mononuclear cell activation: a neuroendocrine-immune interface. Circ Res 93(10), e124-35 (2003) Andersson U, Andersson J, Lindfors A, Wagner K, Moller G, Heusser CH: Simultaneous production of interleukin 2, interleukin 4 and interferon-gamma by activated human blood lymphocytes. Eur J Immunol 20(7), 1591-6 (1990) Anker SD, Rauchhaus M: Insights into the pathogenesis of chronic heart failure: immune activation and cachexia. Curr Opin Cardiol 14(3), 211-6 (1999) Antonijevic IA, Murck H, Frieboes R, Holsboer F, Steiger A: On the gender differences in sleep-endocrine regulation in young normal humans. Neuroendocrinology 70(4), 280-7 (1999) Arlt W, Hewison M: Hormones and immune function: implications of aging. Aging Cell 3(4), 209-16 (2004) Arora SK: Analysis of intracellular cytokines using flowcytometry. Methods Cell Sci 24(1-3), 37-40 (2002) Ashwell JD, Lu FW, Vacchio MS: Glucocorticoids in T cell development and function*. Annu Rev Immunol 18, 309-45 (2000) Auernhammer CJ, Strasburger CJ: Effects of growth hormone and insulin-like growth factor I on the immune system. Eur J Endocrinol 133(6), 635-45 (1995) 56 Literaturverzeichnis Baeza I, Alvarado C, Ariznavarreta C, Castillo C, Tresguerres JA, De la Fuente M: Effect of growth hormone treatment on lymphocyte functions in old male rats. Neuroimmunomodulation 15(4-6), 279-84 (2008) Ball TM, Anderson D, Minto J, Halonen M: Cortisol circadian rhythms and stress responses in infants at risk of allergic disease. J Allergy Clin Immunol 117(2), 306-11 (2006) Banchereau J, Briere F, Caux C, Davoust J, Lebecque S, Liu YJ, Pulendran B, Palucka K: Immunobiology of dendritic cells. Annu Rev Immunol 18, 767-811 (2000) Barbano G, Cappa F, Prigione I, Pistoia V, Cohen A, Chiesa S, Gusmano R, Perfumo F: Plasma levels of soluble CD30 are increased in children with chronic renal failure and with primary growth deficiency and decrease during treatment with recombination human growth hormone. Nephrol Dial Transplant 16(9), 1807-13 (2001) Belkaid Y, Piccirillo CA, Mendez S, Shevach EM, Sacks DL: CD4+CD25+ regulatory T cells control Leishmania major persistence and immunity. Nature 420(6915), 502-7 (2002) Belvisi MG, Hele DJ: Soft steroids: a new approach to the treatment of inflammatory airways diseases. Pulm Pharmacol Ther 16(6), 321-5 (2003) Benedict C, Dimitrov S, Marshall L, Born J: Sleep enhances serum interleukin-7 concentrations in humans. Brain Behav Immun 21(8), 1058-62 (2007) Berczi I: The role of the growth and lactogenic hormone family in immune function. Neuroimmunomodulation 1(4), 201-16 (1994) Berke G: The binding and lysis of target cells by cytotoxic lymphocytes: molecular and cellular aspects. Annu Rev Immunol 12, 735-73 (1994) Bernabei P, Bosticardo M, Losana G, Regis G, Di Paola F, De Angelis S, Giovarelli M, Novelli F: IGF-1 down-regulates IFN-gamma R2 chain surface expression and desensitizes IFN-gamma/STAT-1 signaling in human T lymphocytes. Blood 102(8), 2933-9 (2003) Besedovsky H, del Rey A, Sorkin E, Dinarello CA: Immunoregulatory feedback between interleukin-1 and glucocorticoid hormones. Science 233(4764), 652-4 (1986) Besedovsky L, Lange T, Born J: Sleep and immune function. Pflugers Arch 463(1), 121-37 (2012) 57 Literaturverzeichnis Black PH: Central nervous system-immune system interactions: psychoneuroendocrinology of stress and its immune consequences. Antimicrob Agents Chemother 38(1), 1-6 (1994) Blotta MH, DeKruyff RH, Umetsu DT: Corticosteroids inhibit IL-12 production in human monocytes and enhance their capacity to induce IL-4 synthesis in CD4+ lymphocytes. J Immunol 158(12), 5589-95 (1997) Bollinger T, Bollinger A, Naujoks J, Lange T, Solbach W: The influence of regulatory T cells and diurnal hormone rhythms on T helper cell activity. Immunology 131(4), 488-500 (2010) Bollinger T, Bollinger A, Skrum L, Dimitrov S, Lange T, Solbach W: Sleep-dependent activity of T cells and regulatory T cells. Clin Exp Immunol 155(2), 231-8 (2009) Born J, Fehm HL: Hypothalamus-pituitary-adrenal activity during human sleep: a coordinating role for the limbic hippocampal system. Exp Clin Endocrinol Diabetes 106(3), 153-63 (1998) Born J, Lange T, Hansen K, Molle M, Fehm HL: Effects of sleep and circadian rhythm on human circulating immune cells. J Immunol 158(9), 4454-64 (1997) Bornstein SR, Rutkowski H, Vrezas I: Endocrinol 215(1-2), 135-41 (2004) Cytokines and steroidogenesis. Mol Cell Borrione P, Grasso L, Pautasso M, Parisi A, Quaranta F, Ciminelli E, Di Gianfrancesco A, Di Luigi L, Pigozzi F: Impact of different concentrations of human recombinant growth hormone on T lymphocytes. Int J Immunopathol Pharmacol 25(1), 87-97 (2012) Bouaziz JD, Yanaba K, Tedder TF: Regulatory B cells as inhibitors of immune responses and inflammation. Immunol Rev 224, 201-14 (2008) Brandenberger G, Follenius M: Influence of timing and intensity of musclar exercise on temporal patterns of plasma cortisol levels. J Clin Endocrinol Metab 40(5), 845-9 (1975) Brandenberger G, Follenius M, Hietter B: Feedback from meal-related peaks determines diurnal changes in cortisol response to exercise. J Clin Endocrinol Metab 54(3), 592-6 (1982) Braun CM, Huang SK, Bashian GG, Kagey-Sobotka A, Lichtenstein LM, Essayan DM: Corticosteroid modulation of human, antigen-specific Th1 and Th2 responses. J 58 Literaturverzeichnis Allergy Clin Immunol 100(3), 400-7 (1997) Bresson JL, Jeay S, Gagnerault MC, Kayser C, Beressi N, Wu Z, Kinet S, Dardenne M, Postel-Vinay MC: Growth hormone (GH) and prolactin receptors in human peripheral blood mononuclear cells: relation with age and GH-binding protein. Endocrinology 140(7), 3203-9 (1999) Brinkmann V, Kristofic C: Regulation by corticosteroids of Th1 and Th2 cytokine production in human CD4+ effector T cells generated from CD45RO- and CD45RO+ subsets. J Immunol 155(7), 3322-8 (1995) Brown M, Wittwer C: Flow cytometry: principles and clinical applications in hematology. Clin Chem 46(8 Pt 2), 1221-9 (2000) Bukowski JF, Warner JF, Dennert G, Welsh RM: Adoptive transfer studies demonstrating the antiviral effect of natural killer cells in vivo. J Exp Med 161(1), 4052 (1985) Byron KA, Varigos G, Wootton A: Hydrocortisone inhibition of human interleukin-4. Immunology 77(4), 624-6 (1992) Carvajal CA, Herrada AA, Castillo CR, Contreras FJ, Stehr CB, Mosso LM, Kalergis AM, Fardella CE: Primary aldosteronism can alter peripheral levels of transforming growth factor beta and tumor necrosis factor alpha. J Endocrinol Invest 32(9), 759-65 (2009) Castinetti F, Brue T, Conte-Devolx B: The use of the glucocorticoid receptor antagonist mifepristone in Cushing's syndrome. Curr Opin Endocrinol Diabetes Obes (2012) Castle S, Uyemura K, Wong W, Modlin R, Effros R: Evidence of enhanced type 2 immune response and impaired upregulation of a type 1 response in frail elderly nursing home residents. Mech Ageing Dev 94(1-3), 7-16 (1997) Castro-Magana M, Maddaiah VT, Collipp PJ, Angulo M: Synergistic effects of growth hormone therapy on plasma levels of 11-deoxycortisol and cortisol in growth hormone-deficient children. J Clin Endocrinol Metab 56(4), 662-7 (1983) Cavaillon JM: Cytokines and macrophages. Biomed Pharmacother 48(10), 445-53 (1994) Cederbom L, Hall H, Ivars F: CD4+CD25+ regulatory T cells down-regulate costimulatory molecules on antigen-presenting cells. Eur J Immunol 30(6), 1538-43 (2000) 59 Literaturverzeichnis Chappel S: Growth hormone in immune reconstitution. J Acquir Immune Defic Syndr Hum Retrovirol 20(5), 423-31 (1999) Charloux A, Gronfier C, Chapotot F, Ehrhart J, Piquard F, Brandenberger G: Sleep deprivation blunts the night time increase in aldosterone release in humans. J Sleep Res 10(1), 27-33 (2001) Chen L, Grabowski KA, Xin JP, Coleman J, Huang Z, Espiritu B, Alkan S, Xie HB, Zhu Y, White FA, Clancy J, Jr., Huang H: IL-4 induces differentiation and expansion of Th2 cytokine-producing eosinophils. J Immunol 172(4), 2059-66 (2004) Clark R: The somatogenic hormones and insulin-like growth factor-1: stimulators of lymphopoiesis and immune function. Endocr Rev 18(2), 157-79 (1997) Cohen JL, Trenado A, Vasey D, Klatzmann D, Salomon BL: CD4(+)CD25(+) immunoregulatory T Cells: new therapeutics for graft-versus-host disease. J Exp Med 196(3), 401-6 (2002) Cohen S, Doyle WJ, Alper CM, Janicki-Deverts D, Turner RB: Sleep habits and susceptibility to the common cold. Arch Intern Med 169(1), 62-7 (2009) Corthay A: How do regulatory T cells work? Scand J Immunol 70(4), 326-36 (2009) Dardenne M, Mello-Coelho V, Gagnerault MC, Postel-Vinay MC: Growth hormone receptors and immunocompetent cells. Ann N Y Acad Sci 840, 510-7 (1998) Davidson JR, Moldofsky H, Lue FA: Growth hormone and cortisol secretion in relation to sleep and wakefulness. J Psychiatry Neurosci 16(2), 96-102 (1991) Davila DR, Brief S, Simon J, Hammer RE, Brinster RL, Kelley KW: Role of growth hormone in regulating T-dependent immune events in aged, nude, and transgenic rodents. J Neurosci Res 18(1), 108-16 (1987) Daynes RA, Araneo BA: Contrasting effects of glucocorticoids on the capacity of T cells to produce the growth factors interleukin 2 and interleukin 4. Eur J Immunol 19(12), 2319-25 (1989) de Boer H, Blok GJ, Popp-Snijders C, Stuurman L, Baxter RC, van der Veen E: Monitoring of growth hormone replacement therapy in adults, based on measurement of serum markers. J Clin Endocrinol Metab 81(4), 1371-7 (1996) De Groote D, Zangerle PF, Gevaert Y, Fassotte MF, Beguin Y, Noizat-Pirenne F, Pirenne J, Gathy R, Lopez M, Dehart I, et al.: Direct stimulation of cytokines (IL-1 60 Literaturverzeichnis beta, TNF-alpha, IL-6, IL-2, IFN-gamma and GM-CSF) in whole blood. I. Comparison with isolated PBMC stimulation. Cytokine 4(3), 239-48 (1992) de Kloet ER, Oitzl MS, Joels M: Functional implications of brain corticosteroid receptor diversity. Cell Mol Neurobiol 13(4), 433-55 (1993) de Kloet ER, Oitzl MS, Joels M: Stress and cognition: are corticosteroids good or bad guys? Trends Neurosci 22(10), 422-6 (1999) De Kloet ER, Vreugdenhil E, Oitzl MS, Joels M: Brain corticosteroid receptor balance in health and disease. Endocr Rev 19(3), 269-301 (1998) Delves PJ, Roitt IM: The immune system. First of two parts. N Engl J Med 343(1), 37-49 (2000a) Delves PJ, Roitt IM: The immune system. Second of two parts. N Engl J Med 343(2), 108-17 (2000b) Dhabhar FS: Stress-induced augmentation of immune function--the role of stress hormones, leukocyte trafficking, and cytokines. Brain Behav Immun 16(6), 785-98 (2002) Dhabhar FS: Enhancing versus suppressive effects of stress on immune function: implications for immunoprotection and immunopathology. Neuroimmunomodulation 16(5), 300-17 (2009) Dhabhar FS, Miller AH, Stein M, McEwen BS, Spencer RL: Diurnal and acute stressinduced changes in distribution of peripheral blood leukocyte subpopulations. Brain Behav Immun 8(1), 66-79 (1994) Dimitrov S, Benedict C, Heutling D, Westermann J, Born J, Lange T: Cortisol and epinephrine control opposing circadian rhythms in T cell subsets. Blood 113(21), 5134-43 (2009) Dimitrov S, Lange T, Benedict C, Nowell MA, Jones SA, Scheller J, Rose-John S, Born J: Sleep enhances IL-6 trans-signaling in humans. Faseb J 20(12), 2174-6 (2006) Dimitrov S, Lange T, Fehm HL, Born J: A regulatory role of prolactin, growth hormone, and corticosteroids for human T-cell production of cytokines. Brain Behav Immun 18(4), 368-74 (2004a) Dimitrov S, Lange T, Nohroudi K, Born J: Number and function of circulating human antigen presenting cells regulated by sleep. Sleep 30(4), 401-11 (2007) 61 Literaturverzeichnis Dimitrov S, Lange T, Tieken S, Fehm HL, Born J: Sleep associated regulation of T helper 1/T helper 2 cytokine balance in humans. Brain Behav Immun 18(4), 341-8 (2004b) Dobbs CM, Feng N, Beck FM, Sheridan JF: Neuroendocrine regulation of cytokine production during experimental influenza viral infection: effects of restraint stressinduced elevation in endogenous corticosterone. J Immunol 157(5), 1870-7 (1996) Doganci A, Eigenbrod T, Krug N, De Sanctis GT, Hausding M, Erpenbeck VJ, Haddad el B, Lehr HA, Schmitt E, Bopp T, Kallen KJ, Herz U, Schmitt S, Luft C, Hecht O, Hohlfeld JM, Ito H, Nishimoto N, Yoshizaki K, Kishimoto T, Rose-John S, Renz H, Neurath MF, Galle PR, Finotto S: The IL-6R alpha chain controls lung CD4+CD25+ Treg development and function during allergic airway inflammation in vivo. J Clin Invest 115(2), 313-25 (2005) Duramad P, McMahon CW, Hubbard A, Eskenazi B, Holland NT: Flow cytometric detection of intracellular TH1/TH2 cytokines using whole blood: validation of immunologic biomarker for use in epidemiologic studies. Cancer Epidemiol Biomarkers Prev 13(9), 1452-8 (2004) Edwards CR, Benediktsson R, Lindsay RS, Seckl JR: 11 beta-Hydroxysteroid dehydrogenases: key enzymes in determining tissue-specific glucocorticoid effects. Steroids 61(4), 263-9 (1996) Elenkov IJ: Glucocorticoids and the Th1/Th2 balance. Ann N Y Acad Sci 1024, 13846 (2004) Elenkov IJ, Chrousos GP: Stress Hormones, Th1/Th2 patterns, Pro/Antiinflammatory Cytokines and Susceptibility to Disease. Trends Endocrinol Metab 10(9), 359-368 (1999) Esquifino AI, Alvarez MP, Cano P, Chacon F, Reyes Toso CF, Cardinali DP: 24-hour pattern of circulating prolactin and growth hormone levels and submaxillary lymph node immune responses in growing male rats subjected to social isolation. Endocrine 25(1), 41-8 (2004) Esteban NV, Loughlin T, Yergey AL, Zawadzki JK, Booth JD, Winterer JC, Loriaux DL: Daily cortisol production rate in man determined by stable isotope dilution/mass spectrometry. J Clin Endocrinol Metab 72(1), 39-45 (1991) Everson CA: Sustained sleep deprivation impairs host defense. Am J Physiol 265(5 Pt 2), R1148-54 (1993) Everson CA: Clinical assessment of blood leukocytes, serum cytokines, and serum 62 Literaturverzeichnis immunoglobulins as responses to sleep deprivation in laboratory rats. Am J Physiol Regul Integr Comp Physiol 289(4), R1054-63 (2005) Fearon DT, Locksley RM: The instructive role of innate immunity in the acquired immune response. Science 272(5258), 50-3 (1996) Fehm HL, Holl R, Steiner K, Klein E, Voigt KH: Evidence for ACTH-unrelated mechanisms in the regulation of cortisol secretion in man. Klin Wochenschr 62(1), 19-24 (1984) Figdor CG, van Kooyk Y, Adema GJ: C-type lectin receptors on dendritic cells and Langerhans cells. Nat Rev Immunol 2(2), 77-84 (2002) Fiorentino DF, Zlotnik A, Mosmann TR, Howard M, O'Garra A: IL-10 inhibits cytokine production by activated macrophages. J Immunol 147(11), 3815-22 (1991a) Fiorentino DF, Zlotnik A, Vieira P, Mosmann TR, Howard M, Moore KW, O'Garra A: IL-10 acts on the antigen-presenting cell to inhibit cytokine production by Th1 cells. J Immunol 146(10), 3444-51 (1991b) Fleshner M, Deak T, Nguyen KT, Watkins LR, Maier SF: Endogenous glucocorticoids play a positive regulatory role in the anti-keyhole limpet hemocyanin in vivo antibody response. J Immunol 166(6), 3813-9 (2001) Follenius M, Brandenberger G, Hietter B: Diurnal cortisol peaks and their relationships to meals. J Clin Endocrinol Metab 55(4), 757-61 (1982) Ford PJ: Immunological techniques: ELISA, flow immunohistochemistry. Methods Mol Biol 666, 327-43 (2010) cytometry, and Forsythe P: The nervous system as a critical regulator of immune responses underlying allergy. Curr Pharm Des 18(16), 2290-304 (2012) Franchimont D, Galon J, Gadina M, Visconti R, Zhou Y, Aringer M, Frucht DM, Chrousos GP, O'Shea JJ: Inhibition of Th1 immune response by glucocorticoids: dexamethasone selectively inhibits IL-12-induced Stat4 phosphorylation in T lymphocytes. J Immunol 164(4), 1768-74 (2000) Franchimont D, Louis E, Dewe W, Martens H, Vrindts-Gevaert Y, De Groote D, Belaiche J, Geenen V: Effects of dexamethasone on the profile of cytokine secretion in human whole blood cell cultures. Regul Pept 73(1), 59-65 (1998) Frank MM, Fries LF: The role of complement in inflammation and phagocytosis. Immunol Today 12(9), 322-6 (1991) 63 Literaturverzeichnis Fraser IP, Koziel H, Ezekowitz RA: The serum mannose-binding protein and the macrophage mannose receptor are pattern recognition molecules that link innate and adaptive immunity. Semin Immunol 10(5), 363-72 (1998) Frey FJ, Odermatt A, Frey BM: Glucocorticoid-mediated mineralocorticoid receptor activation and hypertension. Curr Opin Nephrol Hypertens 13(4), 451-8 (2004) Funder JW: Aldosterone receptors. Adv Exp Med Biol 196, 145-52 (1986) Funder JW: Mineralocorticoids, glucocorticoids, receptors and response elements. Science 259(5098), 1132-3 (1993) Funder JW: Mineralocorticoid receptors and glucocorticoid receptors. Clin Endocrinol (Oxf) 45(6), 651-6 (1996) Funder JW: Glucocorticoid and mineralocorticoid receptors: biology and clinical relevance. Annu Rev Med 48, 231-40 (1997) Funder JW: Aldosterone and mineralocorticoid receptors: orphan questions. Kidney Int 57(4), 1358-63 (2000) Funder JW: Mineralocorticoid receptors: distribution and activation. Heart Fail Rev 10(1), 15-22 (2005) Gajewski TF, Fitch FW: Anti-proliferative effect of IFN-gamma in immune regulation. I. IFN-gamma inhibits the proliferation of Th2 but not Th1 murine helper T lymphocyte clones. J Immunol 140(12), 4245-52 (1988) Gajewski TF, Schell SR, Nau G, Fitch FW: Regulation of T-cell activation: differences among T-cell subsets. Immunol Rev 111, 79-110 (1989) Galdiero M, Donnarumma G, Cipollaro de l'Ero G, Marcatili A, Scarfogliero P, Sommese L: Effect of growth hormone, prolactin and insulin on the release of IL-1 alpha, IFN-gamma and IL-4 by staphylococcal enterotoxin A-stimulated splenocytes. Eur Cytokine Netw 6(3), 187-94 (1995) Galdiero M, Vitiello M, Scarfogliero P, Sommese L: Growth hormone release of interleukin-1 alpha, interferon-gamma and interleukin-4 from murine splenocytes stimulated with staphylococcal protein A, toxic shock syndrome toxin-1 and streptococcal lysin S. Eur Cytokine Netw 8(1), 83-90 (1997) Gallagher TF, Yoshida K, Roffwarg HD, Fukushima DK, Weitzman ED, Hellman L: ACTH and cortisol secretory patterns in man. J Clin Endocrinol Metab 36(6), 1058-68 64 Literaturverzeichnis (1973) Gay J, Kokkotou E, O'Brien M, Pothoulakis C, Karalis KP: Corticotropin-releasing hormone deficiency is associated with reduced local inflammation in a mouse model of experimental colitis. Endocrinology 149(7), 3403-9 (2008) Geenen V, Kecha O, Brilot F, Hansenne I, Renard C, Martens H: Thymic T-cell tolerance of neuroendocrine functions: physiology and pathophysiology. Cell Mol Biol (Noisy-le-grand) 47(1), 179-88 (2001) Gleeson M, Bishop NC: The T cell and NK cell immune response to exercise. Ann Transplant 10(4), 43-8 (2005) Graf T: Immunology: blood lines redrawn. Nature 452(7188), 702-3 (2008) Gudewill S, Pollmacher T, Vedder H, Schreiber W, Fassbender K, Holsboer F: Nocturnal plasma levels of cytokines in healthy men. Eur Arch Psychiatry Clin Neurosci 242(1), 53-6 (1992) Guillemin R: Hypothalamic hormones a.k.a. hypothalamic releasing factors. J Endocrinol 184(1), 11-28 (2005) Han Q, Bagheri N, Bradshaw EM, Hafler DA, Lauffenburger DA, Love JC: Polyfunctional responses by human T cells result from sequential release of cytokines. Proc Natl Acad Sci U S A 109(5), 1607-12 (2012) Hattori N: Expression, regulation and biological actions of growth hormone (GH) and ghrelin in the immune system. Growth Horm IGF Res 19(3), 187-97 (2009) Haus E: Chronobiology in the endocrine system. Adv Drug Deliv Rev 59(9-10), 9851014 (2007) Hench PS, Kendall EC, Slocumb CH, Polley HF: Adrenocortical Hormone in Arthritis : Preliminary Report. Ann Rheum Dis 8(2), 97-104 (1949) Hench PS, Kendall EC, Slocumb CH, Polley HF: Cortisone, its effects on rheumatoid arthritis, rheumatic fever, and certain other conditions. Merck Rep 59(4), 9-14 (1950) Herberth G, Heinrich J, Roder S, Figl A, Weiss M, Diez U, Borte M, Herbarth O, Lehmann I: Reduced IFN-gamma- and enhanced IL-4-producing CD4+ cord blood T cells are associated with a higher risk for atopic dermatitis during the first 2 yr of life. Pediatr Allergy Immunol 21(1 Pt 1), 5-13 (2010) Hermus AR, Pieters GF, Smals AG, Benraad TJ, Kloppenborg PW: Plasma 65 Literaturverzeichnis adrenocorticotropin, cortisol, and aldosterone responses to corticotropin-releasing factor: modulatory effect of basal cortisol levels. J Clin Endocrinol Metab 58(1), 18791 (1984) Holl R, Fehm HL, Voigt KH, Teller W: The "midday surge" in plasma cortisol induced by mental stress. Horm Metab Res 16(3), 158-9 (1984) Holland N, Dong J, Garnett E, Shaikh N, Huen K, Harmatz P, Olive A, Winter HS, Gold BD, Cohen SA, Baldassano RN, Kirschner BS, Heyman MB: Reduced intracellular T-helper 1 interferon-gamma in blood of newly diagnosed children with Crohn's disease and age-related changes in Th1/Th2 cytokine profiles. Pediatr Res 63(3), 257-62 (2008) Horne JA: Sleep loss and "divergent" thinking ability. Sleep 11(6), 528-36 (1988) Houssaint E, Sai P: [Development of the immune system]. Ann Endocrinol (Paris) 49(1), 68-76 (1988) Hubl W, Buchner M, Stahl F, Rohde W: Radioimmunoassay for aldosterone in plasma without chromatography. Endokrinologie 66(3), 292-300 (1975) Ibelgaufts H. (2012, 01.2012). COPE: Horst Ibelgaufts' Cytokines & Cells Online Pathfinder Encyclopaedia from 'COPE: Horst Ibelgaufts' Cytokines & Cells Online Pathfinder Encyclopaedia at www.copewithcytokines.org'.Retrieved 18.05.2012, 2012 Irwin M, Miller C, Gillin JC, Demodena A, Ehlers CL: Polysomnographic and spectral sleep EEG in primary alcoholics: an interaction between alcohol dependence and African-American ethnicity. Alcohol Clin Exp Res 24(9), 1376-84 (2000) Janeway C, Travers P, Walport M, Shlomchik M (2009). Janeway Immunologie. Heidelberg, Berlin, Spektrum Akad. Verlag. Jara LJ, Navarro C, Medina G, Vera-Lastra O, Blanco F: Immune-neuroendocrine interactions and autoimmune diseases. Clin Dev Immunol 13(2-4), 109-23 (2006) Jason J, Larned J: Single-cell cytokine profiles in normal humans: comparison of flow cytometric reagents and stimulation protocols. J Immunol Methods 207(1), 13-22 (1997) Jerrells TR: Immunodeficiency associated with ethanol abuse. Adv Exp Med Biol 288, 229-36 (1991) Joels M, de Kloet ER: Mineralocorticoid and glucocorticoid receptors in the brain. Implications for ion permeability and transmitter systems. Prog Neurobiol 43(1), 1-36 66 Literaturverzeichnis (1994) Jung T, Schauer U, Heusser C, Neumann C, Rieger C: Detection of intracellular cytokines by flow cytometry. J Immunol Methods 159(1-2), 197-207 (1993) Kaiser H, Kley HK: Physiologie der Nebennierenrindenhormone. In: Kaiser H: Cortisoltherapie: Corticoide in Klinik und Praxis. 12-33, Thieme, Stuttgart, Stuttgart, 2002 Kalu E, Bhaskaran S, Thum MY, Vishwanatha R, Croucher C, Sherriff E, Ford B, Bansal AS: Serial estimation of Th1:th2 cytokines profile in women undergoing invitro fertilization-embryo transfer. Am J Reprod Immunol 59(3), 206-11 (2008) Kamezaki Y, Katsuura S, Kuwano Y, Tanahashi T, Rokutan K: Circulating cytokine signatures in healthy medical students exposed to academic examination stress. Psychophysiology (2012) Karalis K, Muglia LJ, Bae D, Hilderbrand H, Majzoub JA: CRH and the immune system. J Neuroimmunol 72(2), 131-6 (1997) Kasakura S: [A role for T-helper type 1 and type 2 cytokines in the pathogenesis of various human diseases]. Rinsho Byori 46(9), 915-21 (1998) Kayser FH, Böttger EC, Zinkernagel RM (2005). Medizinische Mikrobiologie. Stuttgart, Thieme. Kelley KW: Cross-talk between the immune and endocrine systems. J Anim Sci 66(8), 2095-108 (1988) Kelley KW, Weigent DA, Kooijman R: Protein hormones and immunity. Brain Behav Immun 21(4), 384-92 (2007) Kendall EC (1950). The development of cortisone as a therapeutic agent - Nobel Lecture, December 11, 1950: 1-19. Kidd P: Th1/Th2 balance: the hypothesis, its limitations, and implications for health and disease. Altern Med Rev 8(3), 223-46 (2003) Kirchner H, Kruse A, Neustock P (1998). Cytokine und Interferone. Botenstoffe des Immunsystems. Heidelberg, Spektrum Akademischer Verlag Heidelberg. Klecha AJ, Barreiro Arcos ML, Frick L, Genaro AM, Cremaschi G: Immuneendocrine interactions in autoimmune thyroid diseases. Neuroimmunomodulation 15(1), 68-75 (2008) 67 Literaturverzeichnis Köhrle J, Petrides PE: Hypothalamisch-hypophysäres System und Zielgewebe Hypothalamus-Hypophysen-Nebennierenrinden - (Zona fasciculata-)Achse. In: Löffler G, Petrides PE,Heinrich PC: Biochemie und Patohobiochemie. 862-869, Springer Verlag Heidelberg, Heidelberg, 2007a Köhrle J, Petrides PE: Hypothalamisch-hypophysäres System und Zielgewebe - Die Wachstumshormon-IGF-Achse. In: Löffler G, Petrides PE,Heinrich PC: Biochemie und Pathobiochemie. 885-891, Springer Verlag Heidelberg, Heidelberg, 2007b Koo GC, Huang C, Camacho R, Trainor C, Blake JT, Sirotina-Meisher A, Schleim KD, Wu TJ, Cheng K, Nargund R, McKissick G: Immune enhancing effect of a growth hormone secretagogue. J Immunol 166(6), 4195-201 (2001) Krueger JM, Obal F, Jr.: Growth hormone-releasing hormone and interleukin-1 in sleep regulation. Faseb J 7(8), 645-52 (1993) Kruse N, Greif M, Moriabadi NF, Marx L, Toyka KV, Rieckmann P: Variations in cytokine mRNA expression during normal human pregnancy. Clin Exp Immunol 119(2), 317-22 (2000) Kunicka JE, Talle MA, Denhardt GH, Brown M, Prince LA, Goldstein G: Immunosuppression by glucocorticoids: inhibition of production of multiple lymphokines by in vivo administration of dexamethasone. Cell Immunol 149(1), 39-49 (1993) Kwak DJ, Augustine NH, Borges WG, Joyner JL, Green WF, Hill HR: Intracellular and extracellular cytokine production by human mixed mononuclear cells in response to group B streptococci. Infect Immun 68(1), 320-7 (2000) Lange T, Dimitrov S, Bollinger T, Diekelmann S, Born J: Sleep after vaccination boosts immunological memory. J Immunol 187(1), 283-90 (2011) Lange T, Dimitrov S, Born J: Effects of sleep and circadian rhythm on the human immune system. Ann N Y Acad Sci 1193, 48-59 (2010) Lange T, Dimitrov S, Fehm HL, Born J: Sleep-like concentrations of growth hormone and cortisol modulate type1 and type2 in-vitro cytokine production in human T cells. Int Immunopharmacol 6(2), 216-25 (2006a) Lange T, Dimitrov S, Fehm HL, Westermann J, Born J: Shift of monocyte function toward cellular immunity during sleep. Arch Intern Med 166(16), 1695-700 (2006b) Langenkamp A, Messi M, Lanzavecchia A, Sallusto F: Kinetics of dendritic cell 68 Literaturverzeichnis activation: impact on priming of TH1, TH2 and nonpolarized T cells. Nat Immunol 1(4), 311-6 (2000) Levi FA, Canon C, Blum JP, Mechkouri M, Reinberg A, Mathe G: Circadian and/or circahemidian rhythms in nine lymphocyte-related variables from peripheral blood of healthy subjects. J Immunol 134(1), 217-22 (1985) Levi FA, Canon C, Touitou Y, Sulon J, Mechkouri M, Ponsart ED, Touboul JP, Vannetzel JM, Mowzowicz I, Reinberg A, et al.: Circadian rhythms in circulating T lymphocyte subtypes and plasma testosterone, total and free cortisol in five healthy men. Clin Exp Immunol 71(2), 329-35 (1988) Liew FY: T(H)1 and T(H)2 cells: a historical perspective. Nat Rev Immunol 2(1), 5560 (2002) Lippincott-Schwartz J, Glickman J, Donaldson JG, Robbins J, Kreis TE, Seamon KB, Sheetz MP, Klausner RD: Forskolin inhibits and reverses the effects of brefeldin A on Golgi morphology by a cAMP-independent mechanism. J Cell Biol 112(4), 567-77 (1991) Liu ML, Xu G, Xue SR, Zhong XC, Chen GX, Chen ZJ: Plasma levels of Th1/Th2 type cytokine are associated with change of prolactin and GH/IGF-I in hemodialysis patients. Int J Artif Organs 31(4), 303-8 (2008) Lloyd M: Philip Showalter Hench, 1896-1965. Rheumatology (Oxford) 41(5), 582-4 (2002) Long EO: Antigen processing for presentation to CD4+ T cells. New Biol 4(4), 27482 (1992) Longobardi-Givani A (1992). Flow Cytometry - First Principles. New York, Wiley Liss. Lucey DR, Clerici M, Shearer GM: Type 1 and type 2 cytokine dysregulation in human infectious, neoplastic, and inflammatory diseases. Clin Microbiol Rev 9(4), 532-62 (1996) Machura E, Mazur B, Kwiecien J, Karczewska K: Intracellular production of IL-2, IL4, IFN-gamma, and TNF-alpha by peripheral blood CD3+ and CD4+ T cells in children with atopic dermatitis. Eur J Pediatr 166(8), 789-95 (2007) Maggi E, Parronchi P, Manetti R, Simonelli C, Piccinni MP, Rugiu FS, De Carli M, Ricci M, Romagnani S: Reciprocal regulatory effects of IFN-gamma and IL-4 on the in vitro development of human Th1 and Th2 clones. J Immunol 148(7), 2142-7 (1992) 69 Literaturverzeichnis Malarkey WB, Wang J, Cheney C, Glaser R, Nagaraja H: Human lymphocyte growth hormone stimulates interferon gamma production and is inhibited by cortisol and norepinephrine. J Neuroimmunol 123(1-2), 180-7 (2002) Mallmann P, Mallmann R, Nadstawek R: [Circadian rhythm of cellular and humoral immunologic parameters]. Immun Infekt 17(3), 97-9 (1989) Martin TR, Mathison JC, Tobias PS, Leturcq DJ, Moriarty AM, Maunder RJ, Ulevitch RJ: Lipopolysaccharide binding protein enhances the responsiveness of alveolar macrophages to bacterial lipopolysaccharide. Implications for cytokine production in normal and injured lungs. J Clin Invest 90(6), 2209-19 (1992) Matalka KZ, Ali DA: Stress-induced versus preovulatory and pregnancy hormonal levels in modulating cytokine production following whole blood stimulation. Neuroimmunomodulation 12(6), 366-74 (2005) Matsushita T, Yanaba K, Bouaziz JD, Fujimoto M, Tedder TF: Regulatory B cells inhibit EAE initiation in mice while other B cells promote disease progression. J Clin Invest 118(10), 3420-30 (2008) Mauri C, Bosma A: Immune regulatory function of B cells. Annu Rev Immunol 30, 221-41 (2012) Mauri C, Ehrenstein MR: The 'short' history of regulatory B cells. Trends Immunol 29(1), 34-40 (2008) McEwen BS: Sleep deprivation as a neurobiologic and physiologic stressor: Allostasis and allostatic load. Metabolism 55(10 Suppl 2), S20-3 (2006) McEwen BS, Biron CA, Brunson KW, Bulloch K, Chambers WH, Dhabhar FS, Goldfarb RH, Kitson RP, Miller AH, Spencer RL, Weiss JM: The role of adrenocorticoids as modulators of immune function in health and disease: neural, endocrine and immune interactions. Brain Res Brain Res Rev 23(1-2), 79-133 (1997) McGillis JP, Park A, Rubin-Fletter P, Turck C, Dallman MF, Payan DG: Stimulation of rat B-lymphocyte proliferation by corticotropin-releasing factor. J Neurosci Res 23(3), 346-52 (1989) McNerlan SE, Rea IM, Alexander HD: A whole blood method for measurement of intracellular TNF-alpha, IFN-gamma and IL-2 expression in stimulated CD3+ lymphocytes: differences between young and elderly subjects. Exp Gerontol 37(2-3), 227-34 (2002) Meerlo P, Sgoifo A, Suchecki D: Restricted and disrupted sleep: effects on autonomic function, neuroendocrine stress systems and stress responsivity. Sleep 70 Literaturverzeichnis Med Rev 12(3), 197-210 (2008) Mellado M, Llorente M, Rodriguez-Frade JM, Lucas P, Martinez C, del Real G: HIV-1 envelope protein gp120 triggers a Th2 response in mice that shifts to Th1 in the presence of human growth hormone. Vaccine 16(11-12), 1111-5 (1998) Migeon CJ, Tyler FH, Mahoney JP, Florentin AA, Castle H, Bliss EL, Samuels LT: The diurnal variation of plasma levels and urinary excretion on 17hydroxycorticosteroids in normal subjects, night workers and blind subjects. J Clin Endocrinol Metab 16(5), 622-33 (1956) Mihailidou AS, Loan Le TY, Mardini M, Funder JW: Glucocorticoids activate cardiac mineralocorticoid receptors during experimental myocardial infarction. Hypertension 54(6), 1306-12 (2009) Miller AH, Spencer RL, hassett J, Kim C, Rhee R, Ciurea D, Dhabhar F, McEwen B, Stein M: Effects of selective type I and II adrenal steroid agonists on immune cell distribution. Endocrinology 135(5), 1934-44 (1994) Miner JN, Hong MH, Negro-Vilar A: New and improved glucocorticoid receptor ligands. Expert Opin Investig Drugs 14(12), 1527-45 (2005) Miyara M, Sakaguchi S: Natural regulatory T cells: mechanisms of suppression. Trends Mol Med 13(3), 108-16 (2007) Mizoguchi A, Bhan AK: A case for regulatory B cells. J Immunol 176(2), 705-10 (2006) Moser M, Murphy KM: Dendritic cell regulation of TH1-TH2 development. Nat Immunol 1(3), 199-205 (2000) Mosmann TR: Regulation of immune responses by T cells with different cytokine secretion phenotypes: role of a new cytokine, cytokine synthesis inhibitory factor (IL10). Int Arch Allergy Appl Immunol 94(1-4), 110-5 (1991) Mosmann TR, Coffman RL: TH1 and TH2 cells: different patterns of lymphokine secretion lead to different functional properties. Annu Rev Immunol 7, 145-73 (1989) Müller U, Vogel P, Alber G, Schaub G: The innate immune system of mammals and insects. Contrib Microbiol. 15, 21-44 (2008) Munck A, Guyre PM, Holbrook NJ: Physiological functions of glucocorticoids in stress and their relation to pharmacological actions. Endocr Rev 5(1), 25-44 (1984) 71 Literaturverzeichnis Murakami M, Sakamoto A, Bender J, Kappler J, Marrack P: CD25+CD4+ T cells contribute to the control of memory CD8+ T cells. Proc Natl Acad Sci U S A 99(13), 8832-7 (2002) Mustafa A, Nyberg F, Mustafa M, Bakhiet M, Mustafa E, Winblad B, Adem A: Growth hormone stimulates production of interferon-gamma by human peripheral mononuclear cells. Horm Res 48(1), 11-5 (1997) Mutschler E, Geisslinger G, Kroemer H, Ruth P, Schäfer-Korting M (2008). Mutschler Arzneimittelwirkungen: Lehrbuch der Pharmakologie und Toxikologie Stuttgart, Wissenschaftliche Verlagsgesellschaft. Myint AM, Leonard BE, Steinbusch HW, Kim YK: Th1, Th2, and Th3 cytokine alterations in major depression. J Affect Disord 88(2), 167-73 (2005) Neumann J: Das Immunsystem In: Neumann J: Immunbiologie Eine Einführung. 1289, Springer Berlin Heidelberg, Berlin, 2008 Nichols CT, Tyler FH: Diurnal variation in adrenal cortical function. Annu Rev Med 18, 313-24 (1967) Noh G, Lee JH: Regulatory B cells and allergic diseases. Allergy Asthma Immunol Res 3(3), 168-77 (2011) Nyberg F: Growth hormone in the brain: characteristics of specific brain targets for the hormone and their functional significance. Front Neuroendocrinol 21(4), 330-48 (2000) Nylander S, Kalies I: Brefeldin A, but not monensin, completely blocks CD69 expression on mouse lymphocytes: efficacy of inhibitors of protein secretion in protocols for intracellular cytokine staining by flow cytometry. J Immunol Methods 224(1-2), 69-76 (1999) Onizuka S, Tawara I, Shimizu J, Sakaguchi S, Fujita T, Nakayama E: Tumor rejection by in vivo administration of anti-CD25 (interleukin-2 receptor alpha) monoclonal antibody. Cancer Res 59(13), 3128-33 (1999) Pala P, Hussell T, Openshaw PJ: Flow cytometric measurement of intracellular cytokines. J Immunol Methods 243(1-2), 107-24 (2000) Palm S, Postler E, Hinrichsen H, Maier H, Zabel P, Kirch W: Twenty-four-hour analysis of lymphocyte subpopulations and cytokines in healthy subjects. Chronobiol Int 13(6), 423-34 (1996) 72 Literaturverzeichnis Pandiyan P, Zheng L, Ishihara S, Reed J, Lenardo MJ: CD4+CD25+Foxp3+ regulatory T cells induce cytokine deprivation-mediated apoptosis of effector CD4+ T cells. Nat Immunol 8(12), 1353-62 (2007) Pasare C, Medzhitov R: Toll pathway-dependent blockade of CD4+CD25+ T cellmediated suppression by dendritic cells. Science 299(5609), 1033-6 (2003) Paul WE, Seder RA: (1994) Lymphocyte responses and cytokines. Cell 76(2), 241-51 Perkoff GT, Eik-Nes K, Nugent CA, Fred HL, Nimer RA, Rush L, Samuels LT, Tyler FH: Studies of the diurnal variation of plasma 17-hydroxycorticosteroids in man. J Clin Endocrinol Metab 19(4), 432-43 (1959) Petersen CM, Christensen EI, Andresen BS, Moller BK: Internalization, lysosomal degradation and new synthesis of surface membrane CD4 in phorbol ester-activated T-lymphocytes and U-937 cells. Exp Cell Res 201(1), 160-73 (1992) Petrovsky N, Harrison LC: Diurnal rhythmicity of human cytokine production: a dynamic disequilibrium in T helper cell type 1/T helper cell type 2 balance? J Immunol 158(11), 5163-8 (1997) Petrovsky N, Harrison LC: The chronobiology of human cytokine production. Int Rev Immunol 16(5-6), 635-49 (1998) Petrovsky N, McNair P, Harrison LC: Diurnal rhythms of pro-inflammatory cytokines: regulation by plasma cortisol and therapeutic implications. Cytokine 10(4), 307-12 (1998) Piccinni MP, Romagnani S: Regulation of fetal allograft survival by a hormonecontrolled Th1- and Th2-type cytokines. Immunol Res 15(2), 141-50 (1996) Piccinni MP, Scaletti C, Maggi E, Romagnani S: Role of hormone-controlled Th1and Th2-type cytokines in successful pregnancy. J Neuroimmunol 109(1), 30-3 (2000) Picker LJ, Singh MK, Zdraveski Z, Treer JR, Waldrop SL, Bergstresser PR, Maino VC: Direct demonstration of cytokine synthesis heterogeneity among human memory/effector T cells by flow cytometry. Blood 86(4), 1408-19 (1995) Pinto RA, Arredondo SM, Bono MR, Gaggero AA, Diaz PV: T helper 1/T helper 2 cytokine imbalance in respiratory syncytial virus infection is associated with increased endogenous plasma cortisol. Pediatrics 117(5), e878-86 (2006) 73 Literaturverzeichnis Postel-Vinay MC, de Mello Coelho V, Gagnerault MC, Dardenne M: Growth hormone stimulates the proliferation of activated mouse T lymphocytes. Endocrinology 138(5), 1816-20 (1997) Prussin C: Cytokine flow cytometry: understanding cytokine biology at the single-cell level. J Clin Immunol 17(3), 195-204 (1997) Pugeat MM, Dunn JF, Nisula BC: Transport of steroid hormones: interaction of 70 drugs with testosterone-binding globulin and corticosteroid-binding globulin in human plasma. J Clin Endocrinol Metab 53(1), 69-75 (1981) Rambach G, Wurzner R, Speth C: Complement: an efficient sword of innate immunity. Contrib Microbiol 15, 78-100 (2008) Ramirez F: Glucocorticoids induce a Th2 response in vitro. Dev Immunol 6(3-4), 233-43 (1998) Ramirez F, Fowell DJ, Puklavec M, Simmonds S, Mason D: Glucocorticoids promote a TH2 cytokine response by CD4+ T cells in vitro. J Immunol 156(7), 2406-12 (1996) Ramos SB, Brenu EW, Christy R, Gray B, McNaughton L, Tajouri L, Van Driel M, Marshall-Gradisnik SM: Assessment of immune function after short-term administration of recombinant human growth hormone in healthy young males. Eur J Appl Physiol 111(7), 1307-12 (2011) Rechtschaffen A, Kahles A: A manual of standardized terminology, techniques, and scoring system for sleep stages of human subjects. In: Berger JR, Dement WC,Jacobson A: National Institutes of Health Publication No. 204. US Government Printing Office, Washington DC Washington DC, 1968 Redwine L, Dang J, Hall M, Irwin M: Disordered sleep, nocturnal cytokines, and immunity in alcoholics. Psychosom Med 65(1), 75-85 (2003) Ricklin D, Hajishengallis G, Yang K, Lambris JD: Complement: a key system for immune surveillance and homeostasis. Nat Immunol 11(9), 785-97 (2010) Rimmele U, Meier F, Lange T, Born J: Suppressing the morning rise in cortisol impairs free recall. Learn Mem 17(4), 186-90 (2010) Rink L, Cakman I, Kirchner H: Altered cytokine production in the elderly. Mech Ageing Dev 102(2-3), 199-209 (1998) Rivier C, Vale W: Modulation of stress-induced ACTH release by corticotropinreleasing factor, catecholamines and vasopressin. Nature 305(5932), 325-7 (1983) 74 Literaturverzeichnis Romagnani S: Type 1 T helper and type 2 T helper cells: functions, regulation and role in protection and disease. Int J Clin Lab Res 21(2), 152-8 (1991) Rostaing L, Tkaczuk J, Durand M, Peres C, Durand D, de Preval C, Ohayon E, Abbal M: Kinetics of intracytoplasmic Th1 and Th2 cytokine production assessed by flow cytometry following in vitro activation of peripheral blood mononuclear cells. Cytometry 35(4), 318-28 (1999) Rupprecht R, Arriza JL, Spengler D, Reul JM, Evans RM, Holsboer F, Damm K: Transactivation and synergistic properties of the mineralocorticoid receptor: relationship to the glucocorticoid receptor. Mol Endocrinol 7(4), 597-603 (1993) Sack U, Rothe G, Barlage S, Gruber R, Kabelitz D, Kleine TO, Lun A, Renz H, Ruf A, Schmitz G: Durchflußzytometrie in der Klinischen Diagnostik. LaboratoriumsMedizin / Journal of Laboratory Medicine 24(6-7), 277-297 (2000) Sad S, Mosmann TR: Single IL-2-secreting precursor CD4 T cell can develop into either Th1 or Th2 cytokine secretion phenotype. J Immunol 153(8), 3514-22 (1994) Sakaguchi S: Naturally arising CD4+ regulatory t cells for immunologic self-tolerance and negative control of immune responses. Annu Rev Immunol 22, 531-62 (2004) Sakaguchi S: Naturally arising Foxp3-expressing CD25+CD4+ regulatory T cells in immunological tolerance to self and non-self. Nat Immunol 6(4), 345-52 (2005) Sakaguchi S: Regulatory T cells: history and perspective. Methods Mol Biol 707, 317 (2011) Sapolsky R, Rivier C, Yamamoto G, Plotsky P, Vale W: Interleukin-1 stimulates the secretion of hypothalamic corticotropin-releasing factor. Science 238(4826), 522-4 (1987) Sauer J, Castren M, Hopfner U, Holsboer F, Stalla GK, Arzt E: Inhibition of lipopolysaccharide-induced monocyte interleukin-1 receptor antagonist synthesis by cortisol: involvement of the mineralocorticoid receptor. J Clin Endocrinol Metab 81(1), 73-9 (1996) Schiebler TH: Histologie, Gewebelehre - Knochen. In: Schiebler TH: Anatomie. 4953, Springer Medizin Verlag Heidelberg, Heidelberg, 2005 Schreiber AM: Visualizing and quantifying the suppressive effects of glucocorticoids on the tadpole immune system in vivo. Adv Physiol Educ 35(4), 445-53 (2011) 75 Literaturverzeichnis Schubert C, Schussler G: [Psychoneuroimmunology: an update]. Z Psychosom Med Psychother 55(1), 3-26 (2009) Schuerwegh AJ, De Clerck LS, Bridts CH, Stevens WJ: Comparison of intracellular cytokine production with extracellular cytokine levels using two flow cytometric techniques. Cytometry B Clin Cytom 55(1), 52-8 (2003) Schuerwegh AJ, Stevens WJ, Bridts CH, De Clerck LS: Evaluation of monensin and brefeldin A for flow cytometric determination of interleukin-1 beta, interleukin-6, and tumor necrosis factor-alpha in monocytes. Cytometry 46(3), 172-6 (2001) Schultz C, Rott C, Temming P, von Puttkammer J, Bucsky P: Influence of specimen age and use of different negative controls in determination of intracytoplasmic levels of cytokines after whole-blood culture assay. Clin Diagn Lab Immunol 9(2), 295-8 (2002) Schwab R, Russo C, Weksler ME: Altered major histocompatibility complexrestricted antigen recognition by T cells from elderly humans. Eur J Immunol 22(11), 2989-93 (1992) Schwarz MJ, Chiang S, Muller N, Ackenheil M: T-helper-1 and T-helper-2 responses in psychiatric disorders. Brain Behav Immun 15(4), 340-70 (2001) Shevach EM: Regulatory T cells in autoimmmunity*. Annu Rev Immunol 18, 423-49 (2000) Siejka A, Lawnicka H, Komorowski J, Stepien T, Krupinski R, Stepien H: Effect of growth hormone-releasing hormone (GHRH) and GHRH antagonist (MZ-4-71) on interferon-gamma secretion from human peripheral blood mononuclear cells in vitro. Neuropeptides 38(1), 35-9 (2004) Siejka A, Stepien T, Lawnicka H, Krupinski R, Komorowski J, Stepien H: Effect of the growth hormone-releasing hormone [GHRH(1-44)NH2] on IL-6 and IL-8 secretion from human peripheral blood mononuclear cells in vitro. Endocr Regul 39(1), 7-11 (2005a) Siejka A, Stepien T, Lawnicka H, Krupinski R, Komorowski J, Stepien H: [Evaluation of the effect of GHRH(1-44)NH2 on the secretion of interleukin-2 (IL-2) and soluble IL-2 receptor alpha (sIL-2Ralpha) from human peripheral blood mononuclear cells in vitro]. Endokrynol Pol 56(5), 773-8 (2005b) Singh VK: Stimulatory effect of corticotropin-releasing neurohormone on human lymphocyte proliferation and interleukin-2 receptor expression. J Neuroimmunol 23(3), 257-62 (1989) 76 Literaturverzeichnis Skapenko A, Schulze-Koops H: Analysis of Th1/Th2 T-cell subsets. Methods Mol Med 136, 87-96 (2007) Skjolaas KA, Grieger DM, Hill CM, Minton JE: Glucocorticoid regulation of type 1 and type 2 cytokines in cultured porcine splenocytes. Vet Immunol Immunopathol 87(1-2), 79-87 (2002) Skjolaas KA, Minton JE: Does cortisol bias cytokine production in cultured porcine splenocytes to a Th2 phenotype? Vet Immunol Immunopathol 87(3-4), 451-8 (2002) Snijdewint FG, Kapsenberg ML, Wauben-Penris PJ, Bos JD: Corticosteroids classdependently inhibit in vitro Th1- and Th2-type cytokine production. Immunopharmacology 29(2), 93-101 (1995) Sommese L, Donnarumma G, de l'Ero C, Marcatili A, Vitiello M, Galdiero M: Growth hormone modulates IL-alpha and IFN-gamma release by murine splenocytes activated by LPS or porins of Salmonella typhimurium. J Med Microbiol 45(1), 40-7 (1996) Song C, Halbreich U, Han C, Leonard BE, Luo H: Imbalance between pro- and antiinflammatory cytokines, and between Th1 and Th2 cytokines in depressed patients: the effect of electroacupuncture or fluoxetine treatment. Pharmacopsychiatry 42(5), 182-8 (2009) Spencer RL, Young EA, Choo PH, McEwen BS: Adrenal steroid type I and type II receptor binding: estimates of in vivo receptor number, occupancy, and activation with varying level of steroid. Brain Res 514(1), 37-48 (1990) Steinman RM, Hemmi H: Dendritic cells: translating innate to adaptive immunity. Curr Top Microbiol Immunol 311, 17-58 (2006) Stephens LA, Mottet C, Mason D, Powrie F: Human CD4(+)CD25(+) thymocytes and peripheral T cells have immune suppressive activity in vitro. Eur J Immunol 31(4), 1247-54 (2001) Stepien T, Lawnicka H, Komorowski J, Stepien H, Siejka A: Growth hormonereleasing hormone stimulates the secretion of interleukin 17 from human peripheral blood mononuclear cells in vitro. Neuro Endocrinol Lett 31(6), 852-6 (2010) Sternberg EM: Neuroendocrine regulation of autoimmune/inflammatory disease. J Endocrinol 169(3), 429-35 (2001) Stevanovic S: Structural basis of immunogenicity. Transpl Immunol 10(2-3), 133-6 (2002) 77 Literaturverzeichnis Stroopinsky D, Avivi I, Rowe JM, Avigan D, Katz T: Allogeneic induced human FOXP3(+)IFN-gamma(+) T cells exhibit selective suppressive capacity. Eur J Immunol 39(10), 2703-15 (2009) Suda T, Tomori N, Yajima F, Sumitomo T, Nakagami Y, Ushiyama T, Demura H, Shizume K: Immunoreactive corticotropin-releasing factor in human plasma. J Clin Invest 76(5), 2026-9 (1985) Suda T, Tomori N, Yajima F, Ushiyama T, Sumitomo T, Nakagami Y, Demura H, Shizume K: A short negative feedback mechanism regulating corticotropin-releasing hormone release. J Clin Endocrinol Metab 64(5), 909-13 (1987) Suda T, Yajima F, Tomori N, Sumitomo T, Nakagami Y, Ushiyama T, Demura H, Shizume K: Inhibitory effect of adrenocorticotropin on corticotropin- releasing factor release from rat hypothalamus in vitro. Endocrinology 118(1), 459-61 (1986) Suzuki H, Zelphati O, Hildebrand G, Leserman L: CD4 and CD7 molecules as targets for drug delivery from antibody bearing liposomes. Exp Cell Res 193(1), 1129 (1991) Svedmyr E, Jondal M: Cytotoxic effector cells specific for B Cell lines transformed by Epstein-Barr virus are present in patients with infectious mononucleosis. Proc Natl Acad Sci U S A 72(4), 1622-6 (1975) Swain SL, Weinberg AD, English M, Huston G: IL-4 directs the development of Th2like helper effectors. J Immunol 145(11), 3796-806 (1990) Szczypka M, Ploch S, Obminska-Mrukowicz B: Modulation of Th1/Th2 cytokine production by selective and nonselective phosphodiesterase inhibitors administered to mice. Pharmacol Rep 64(1), 179-84 (2012) Takagi K, Suzuki F, Barrow RE, Wolf SE, Herndon DN: Recombinant human growth hormone modulates Th1 and Th2 cytokine response in burned mice. Ann Surg 228(1), 106-11 (1998) Takato-Kaji R, Totsuka M, Ise W, Nishikawa M, Hachimura S, Kaminogawa S: T-cell receptor antagonist modifies cytokine secretion profile of naive CD4+ T cells and their differentiation into type-1 and type-2 helper T cells. Immunol Lett 96(1), 39-45 (2005) Tanaka J, Fujita H, Matsuda S, Toku K, Sakanaka M, Maeda N: Glucocorticoid- and mineralocorticoid receptors in microglial cells: the two receptors mediate differential effects of corticosteroids. Glia 20(1), 23-37 (1997) 78 Literaturverzeichnis Tang Q, Bluestone JA: The Foxp3+ regulatory T cell: a jack of all trades, master of regulation. Nat Immunol 9(3), 239-44 (2008) Tayebi H, Lienard A, Billot M, Tiberghien P, Herve P, Robinet E: Detection of intracellular cytokines in citrated whole blood or marrow samples by flow cytometry. J Immunol Methods 229(1-2), 121-30 (1999) Thornton AM, Shevach EM: CD4+CD25+ immunoregulatory T cells suppress polyclonal T cell activation in vitro by inhibiting interleukin 2 production. J Exp Med 188(2), 287-96 (1998) Tosi MF: Innate immune responses to infection. J Allergy Clin Immunol 116(2), 2419; quiz 250 (2005) Tripathi A, Sodhi A: Growth hormone-induced production of cytokines in murine peritoneal macrophages in vitro: role of JAK/STAT, PI3K, PKC and MAP kinases. Immunobiology 214(6), 430-40 (2009) Tu W, Cheung PT, Lau YL: IGF-I increases interferon-gamma and IL-6 mRNA expression and protein production in neonatal mononuclear cells. Pediatr Res 46(6), 748-54 (1999) Tyrrell JB, Findling JW, Aron DC, Fitzgerald PA, Forsham PH: An overnight highdose dexamethasone suppression test for rapid differential diagnosis of Cushing's syndrome. Ann Intern Med 104(2), 180-6 (1986) Valtorta A, Moretta A, Maccario R, Bozzola M, Severi F: Influence of growth hormone-releasing hormone (GHRH) on phytohemagglutinin-induced lymphocyte activation: comparison of two synthetic forms. GHRH and PHA-induced lymphocyte activation. Thymus 18(1), 51-9 (1991) Van Cauter E, Copinschi G: Interrelationships between growth hormone and sleep. Growth Horm IGF Res 10 Suppl B, S57-62 (2000) Van Cauter E, Leproult R, Plat L: Age-related changes in slow wave sleep and REM sleep and relationship with growth hormone and cortisol levels in healthy men. Jama 284(7), 861-8 (2000) Vedder H, Lanquillon S, Krieg JC (2003). The mineralocorticoid receptor system and tumor-necrosis factor-alpha secretion in major depression. The Third German Endocrine Brain Immune Network. München. Veldhuis JD, Iranmanesh A, Weltman A: Elements in the pathophysiology of diminished growth hormone (GH) secretion in aging humans. Endocrine 7(1), 41-8 (1997) 79 Literaturverzeichnis Villadangos JA, Schnorrer P: Intrinsic and cooperative antigen-presenting functions of dendritic-cell subsets in vivo. Nat Rev Immunol 7(7), 543-55 (2007) Visser J, van Boxel-Dezaire A, Methorst D, Brunt T, de Kloet ER, Nagelkerken L: Differential regulation of interleukin-10 (IL-10) and IL-12 by glucocorticoids in vitro. Blood 91(11), 4255-64 (1998) von Andrian UH, Mackay CR: T-cell function and migration. Two sides of the same coin. N Engl J Med 343(14), 1020-34 (2000) Waldrop SL, Pitcher CJ, Peterson DM, Maino VC, Picker LJ: Determination of antigen-specific memory/effector CD4+ T cell frequencies by flow cytometry: evidence for a novel, antigen-specific homeostatic mechanism in HIV-associated immunodeficiency. J Clin Invest 99(7), 1739-50 (1997) Weitzman ED: Biologic rhythms and hormone secretion patterns. Hosp Pract 11(8), 79-86 (1976a) Weitzman ED: Circadian rhythms and episodic hormone secretion in man. Annu Rev Med 27, 225-43 (1976b) Weitzman ED, Czeisler CA, Zimmerman JC, Moore-Ede MC: Biological rhythms in man: relationship of sleep-wake, cortisol, growth hormone, and temperature during temporal isolation. Adv Biochem Psychopharmacol 28, 475-99 (1981) White PC, Mune T, Agarwal AK: 11 beta-Hydroxysteroid dehydrogenase and the syndrome of apparent mineralocorticoid excess. Endocr Rev 18(1), 135-56 (1997) Whitesman S, Booth R: Psychoneuroimmunology--mind-brain-immune interactions. S Afr Med J 94(4), 259-61 (2004) Wu CY, Sarfati M, Heusser C, Fournier S, Rubio-Trujillo M, Peleman R, Delespesse G: Glucocorticoids increase the synthesis of immunoglobulin E by interleukin 4stimulated human lymphocytes. J Clin Invest 87(3), 870-7 (1991) Yanaba K: [Regulatory B cells]. Nihon Rinsho Meneki Gakkai Kaishi 32(3), 135-41 (2009) Yang HZ, Li Z, Liu HZ, Mi S, Hu ZW: [The identification and advances of regulatory B cells]. Sheng Li Ke Xue Jin Zhan 40(4), 297-302 (2009) Yaqoob P, Newsholme EA, Calder PC: Comparison of cytokine production in cultures of whole human blood and purified mononuclear cells. Cytokine 11(8), 600-5 80 Literaturverzeichnis (1999) Yeager MP, Pioli PA, Wardwell K, Beach ML, Martel P, Lee HK, Rassias AJ, Guyre PM: In vivo exposure to high or low cortisol has biphasic effects on inflammatory response pathways of human monocytes. Anesth Analg 107(5), 1726-34 (2008) Yssel H, De Waal Malefyt R, Roncarolo MG, Abrams JS, Lahesmaa R, Spits H, de Vries JE: IL-10 is produced by subsets of human CD4+ T cell clones and peripheral blood T cells. J Immunol 149(7), 2378-84 (1992) Zabel P, Linnemann K, Schlaak M: [Circadian rhythm in cytokines]. Immun Infekt 21 Suppl 1, 38-40 (1993) Zikorsky B, Lange T, Born J: Function of Sleep in Human Adaptive Immunity. In: Sperner-Unterweger B, Fleischhacker WW,Kaschka WP: Psychoneuroimmunology. Hypotheses and Current Research. 20, 7-19, K, Basel, 2001 Zinkernagel RM, Doherty PC: Immunological surveillance against altered self components by sensitised T lymphocytes in lymphocytic choriomeningitis. Nature 251(5475), 547-8 (1974a) Zinkernagel RM, Doherty PC: Restriction of in vitro T cell-mediated cytotoxicity in lymphocytic choriomeningitis within a syngeneic or semiallogeneic system. Nature 248(450), 701-2 (1974b) 81 Anhang 7. Anhang 82 Anhang 83 Danksagung 8. Danksagung An erster Stelle möchte ich meinem Doktorvater, Herrn Professor Dr. med. J. Born für die Überlassung des Promotionsthemas herzlich danken. Insbesondere bedanke ich mich für das nochmals entgegenbrachte Vertrauen, mit dieser Arbeit promovieren zu dürfen. Zu tiefem Dank bin ich Frau Dr. med. T. Lange und Herrn Dr. Dipl.- Biol. S. Dimitrov für die fürsorgliche Betreuung im Rahmen des experimentellen Abschnittes verpflichtet. Ihre Geduld und das Engagement in der langen Erstellungsphase dieser Promotionsarbeit möchte ich an dieser Stelle hervorheben. Mein besonderer Dank gilt Professor Dr. med. Dr. med. dent. P. Sieg, Direktor der Klinik für Kiefer- und Gesichtschirurgie sowie meinen Kollegen, die mich in der Zeit meines Zweitstudiums und während der Fertigstellung dieser Arbeit entlasteten. Ferner bedanke ich mich bei Frau C. Zinke und A. Otterbein für die technische Beratung während der Durchführung des labormedizinischen Teils der Arbeit. Herrn Privatdozent Dr. Dipl.-Ing. P. D. Sindilariu bin ich für die kompetente Beratung und Hilfe bei der statistischen Auswertung sehr verbunden. Ich danke auch meiner Kondoktorandin und Freundin Frau W. Becker für die wunderbare Zusammenarbeit im einsamen Labor. Nicht zuletzt möchte ich Dir Attila für die konstruktive Kritik bei der Überarbeitung des Manuskriptes, Deine liebevolle Unterstützung und dein Verständnis in den vergangenen Jahren danken. Ohne Deine nahezu endlose Geduld hätte ich diese Arbeit wohl nicht mehr fertig gestellt. 84 Lebenslauf 9. Lebenslauf Persönliche Daten Name: Wolf Vorname: Melanie Geburtsdatum: 26.09.1977 Geburtsort: Neubrandenburg Staatsangehörigkeit: deutsch Familienstand: ledig Schulausbildung und beruflicher Werdegang 1984 -1989 Gesamtschule Polytechnische Oberschule Dessau 1989 -1997 Fürst Franz Gymnasium Dessau 1997 Abitur 10/1997-10/2005 Studium der Humanmedizin am Universitätsklinikum Schleswig-Holstein, Campus Lübeck 10/2005 Approbation als Ärztin seit 11/2005 Tätigkeit als wissenschaftliche Assistentin in der Klinik für Kiefer- und Gesichtschirurgie am Universitätsklinikum Schleswig-Holstein, Campus Lübeck 10/2006 -11/2011 Studium der Zahnmedizin an der Universität Hamburg und an der CAU Kiel 11/2011 Approbation als Zahnärztin Lübeck, den 28.06.2012 85 Veröffentlichung der Arbeitsergebnisse 10. Veröffentlichung der Arbeitsergebnisse Dimitrov S, Lange T, Fehm HL, Born J: A regulatory role of prolactin, growth hormone, and corticosteroids for human T-cell production of cytokines. Brain Behav Immun 18(4), 368-74 (2004) 86